

FACT Disrupts Nucleosome Structure by Binding H2A-H2B with Conserved Peptide Motifs
- PMID: 26455391
- PMCID: PMC4620744
- DOI: 10.1016/j.molcel.2015.09.008
FACT Disrupts Nucleosome Structure by Binding H2A-H2B with Conserved Peptide Motifs
Abstract
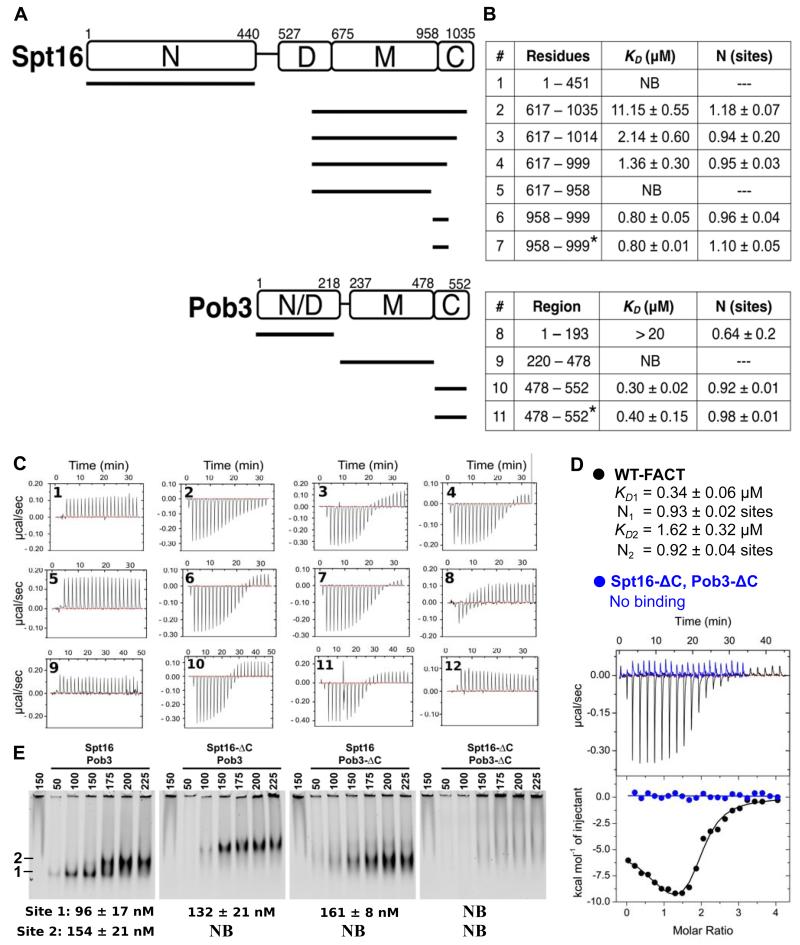
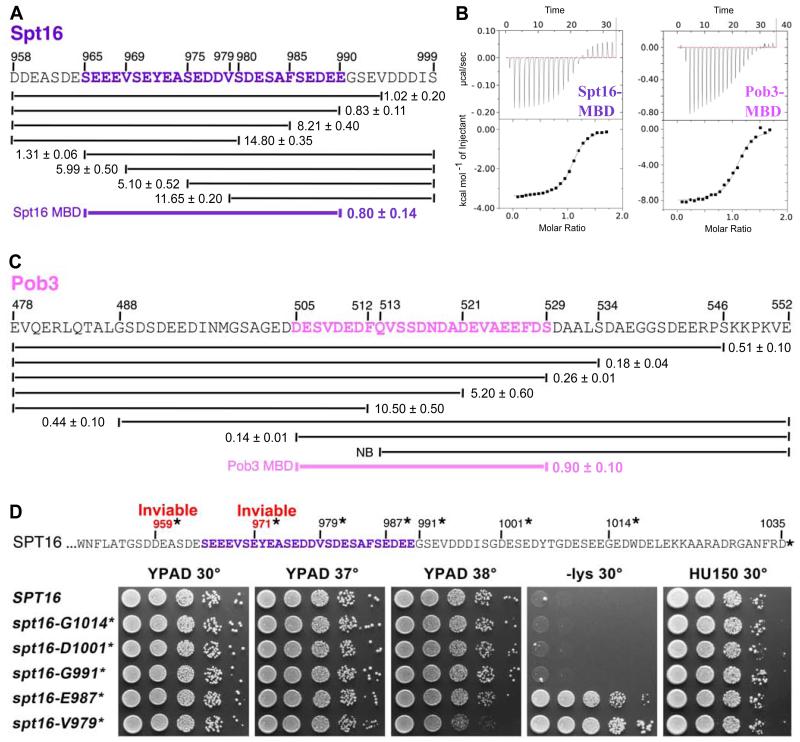
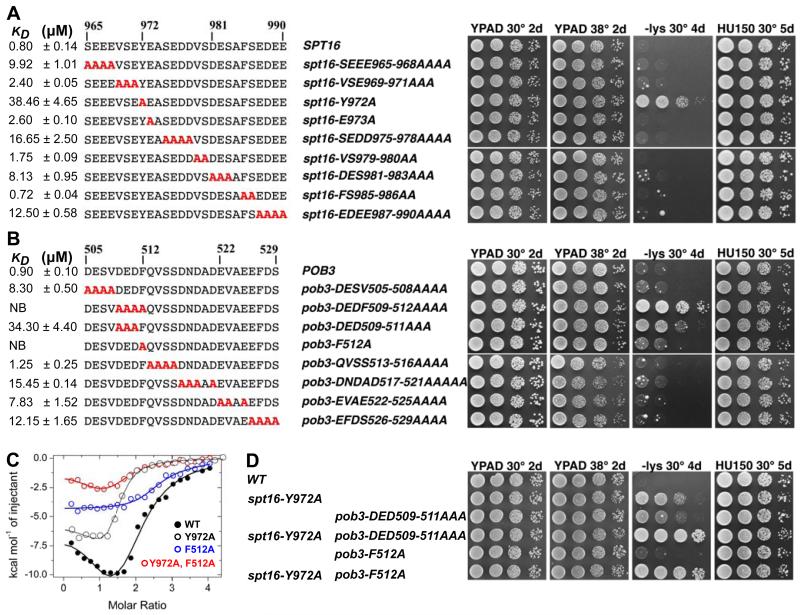
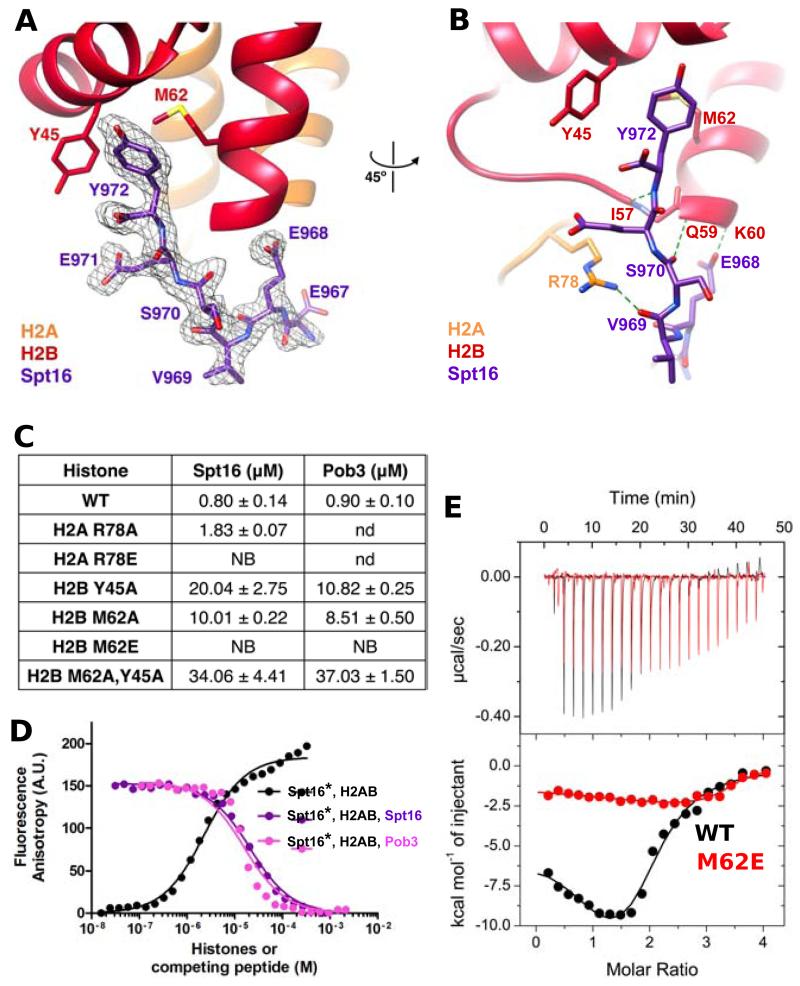
FACT, a heterodimer of Spt16 and Pob3, is an essential histone chaperone. We show that the H2A-H2B binding activity that is central to FACT function resides in short acidic regions near the C termini of each subunit. Mutations throughout these regions affect binding and cause correlated phenotypes that range from mild to lethal, with the largest individual contributions unexpectedly coming from an aromatic residue and a nearby carboxylate residue within each domain. Spt16 and Pob3 bind overlapping sites on H2A-H2B, and Spt16-Pob3 heterodimers simultaneously bind two H2A-H2B dimers, the same stoichiometry as the components of a nucleosome. An Spt16:H2A-H2B crystal structure explains the biochemical and genetic data, provides a model for Pob3 binding, and implies a mechanism for FACT reorganization that we confirm biochemically. Moreover, unexpected similarity to binding of ANP32E and Swr1 with H2A.Z-H2B reveals that diverse H2A-H2B chaperones use common mechanisms of histone binding and regulating nucleosome functions.
Keywords: Spt16-Pob3; crystal structure; histone binding; histone chaperone; nucleosome reorganization.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Residues in the Nucleosome Acidic Patch Regulate Histone Occupancy and Are Important for FACT Binding in Saccharomyces cerevisiae.Genetics. 2017 Jul;206(3):1339-1348. doi: 10.1534/genetics.117.201939. Epub 2017 May 3. Genetics. 2017. PMID: 28468903 Free PMC article.
-
The Abundant Histone Chaperones Spt6 and FACT Collaborate to Assemble, Inspect, and Maintain Chromatin Structure in Saccharomyces cerevisiae.Genetics. 2015 Nov;201(3):1031-45. doi: 10.1534/genetics.115.180794. Epub 2015 Sep 28. Genetics. 2015. PMID: 26416482 Free PMC article.
-
Insight into the mechanism of nucleosome reorganization from histone mutants that suppress defects in the FACT histone chaperone.Genetics. 2011 Aug;188(4):835-46. doi: 10.1534/genetics.111.128769. Epub 2011 May 30. Genetics. 2011. PMID: 21625001 Free PMC article.
-
Nucleosome adaptability conferred by sequence and structural variations in histone H2A-H2B dimers.Curr Opin Struct Biol. 2015 Jun;32:48-57. doi: 10.1016/j.sbi.2015.02.004. Epub 2015 Feb 27. Curr Opin Struct Biol. 2015. PMID: 25731851 Free PMC article. Review.
-
de FACTo nucleosome dynamics.J Biol Chem. 2006 Aug 18;281(33):23297-301. doi: 10.1074/jbc.R600007200. Epub 2006 Jun 9. J Biol Chem. 2006. PMID: 16766522 Review.
Cited by
-
DJ-1 Proteoforms in Breast Cancer Cells: The Escape of Metabolic Epigenetic Misregulation.Cells. 2020 Aug 26;9(9):1968. doi: 10.3390/cells9091968. Cells. 2020. PMID: 32858971 Free PMC article.
-
Establishment and Maintenance of Chromatin Architecture Are Promoted Independently of Transcription by the Histone Chaperone FACT and H3-K56 Acetylation in Saccharomyces cerevisiae.Genetics. 2019 Mar;211(3):877-892. doi: 10.1534/genetics.118.301853. Epub 2019 Jan 24. Genetics. 2019. PMID: 30679261 Free PMC article.
-
FACT and Ubp10 collaborate to modulate H2B deubiquitination and nucleosome dynamics.Elife. 2019 Jan 25;8:e40988. doi: 10.7554/eLife.40988. Elife. 2019. PMID: 30681413 Free PMC article.
-
DNA-dependent RNA polymerases in plants.Plant Cell. 2023 Sep 27;35(10):3641-3661. doi: 10.1093/plcell/koad195. Plant Cell. 2023. PMID: 37453082 Free PMC article.
-
The fork protection complex recruits FACT to reorganize nucleosomes during replication.Nucleic Acids Res. 2022 Feb 22;50(3):1317-1334. doi: 10.1093/nar/gkac005. Nucleic Acids Res. 2022. PMID: 35061899 Free PMC article.
References
-
- Anderson M, Huh JH, Ngo T, Lee A, Hernandez G, Pang J, Perkins J, Dutnall RN. Co-expression as a convenient method for the production and purification of core histones in bacteria. Protein Expr Purif. 2010;72:194–204. - PubMed
-
- Belotserkovskaya R, Oh S, Bondarenko VA, Orphanides G, Studitsky VM, Reinberg D. FACT facilitates transcription-dependent nucleosome alteration. Science. 2003;301:1090–1093. - PubMed
-
- Belotserkovskaya R, Saunders A, Lis JT, Reinberg D. Transcription through chromatin: understanding a complex FACT. Biochim Biophys Acta. 2004;1677:87–99. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
