

Identification of AMPK Phosphorylation Sites Reveals a Network of Proteins Involved in Cell Invasion and Facilitates Large-Scale Substrate Prediction
- PMID: 26456332
- PMCID: PMC4635044
- DOI: 10.1016/j.cmet.2015.09.009
Identification of AMPK Phosphorylation Sites Reveals a Network of Proteins Involved in Cell Invasion and Facilitates Large-Scale Substrate Prediction
Abstract
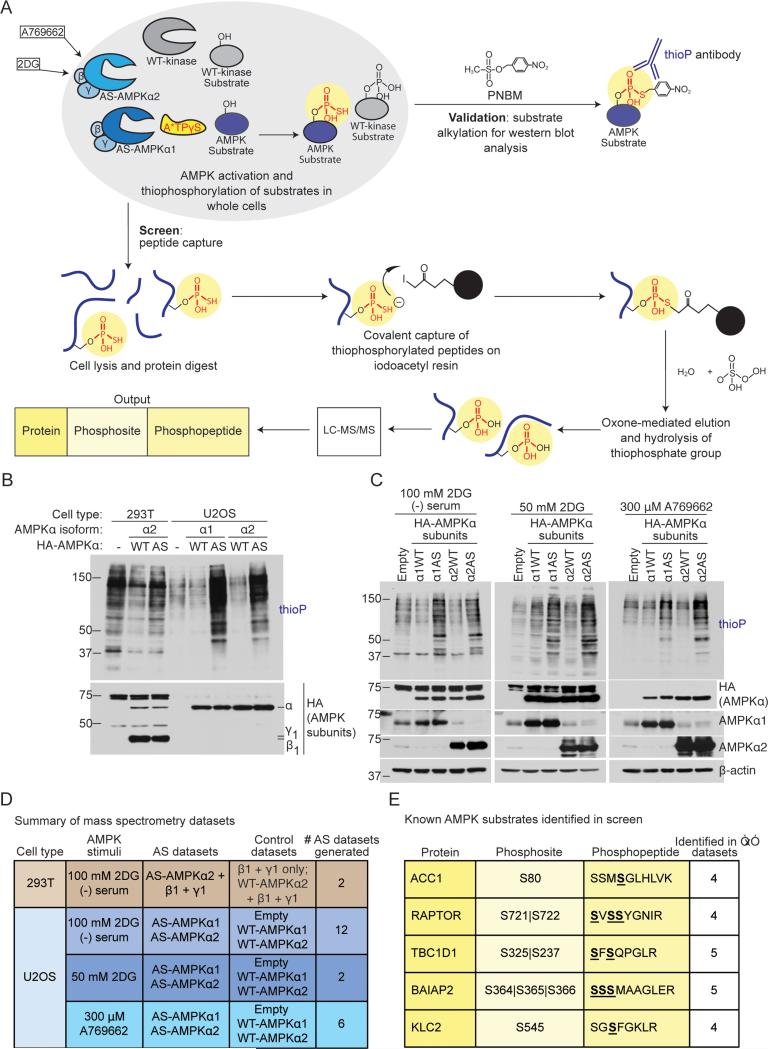
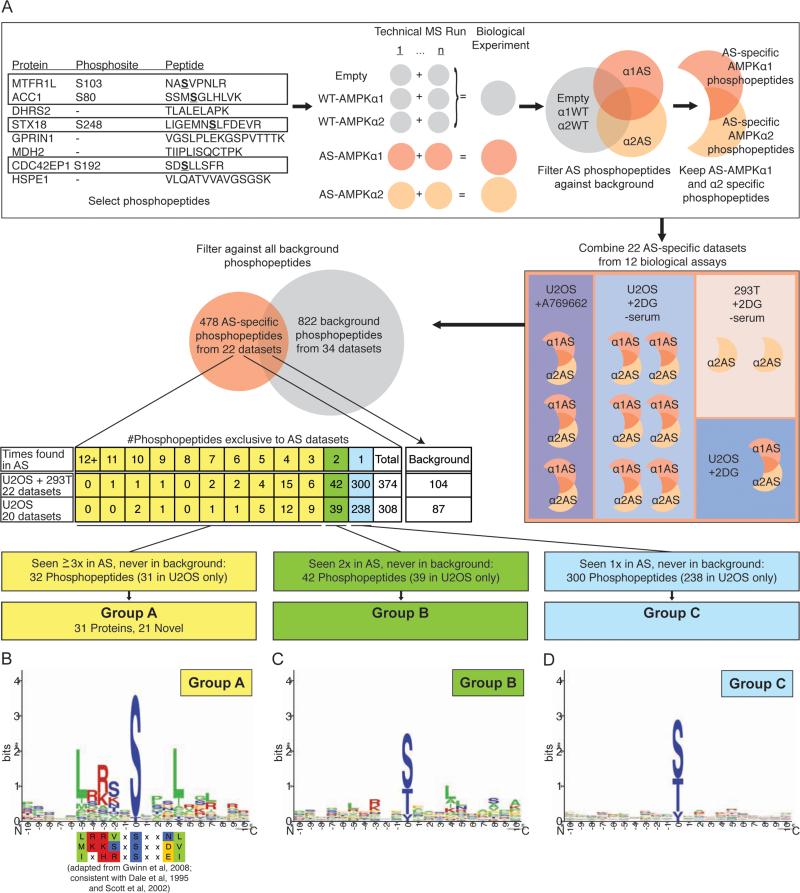
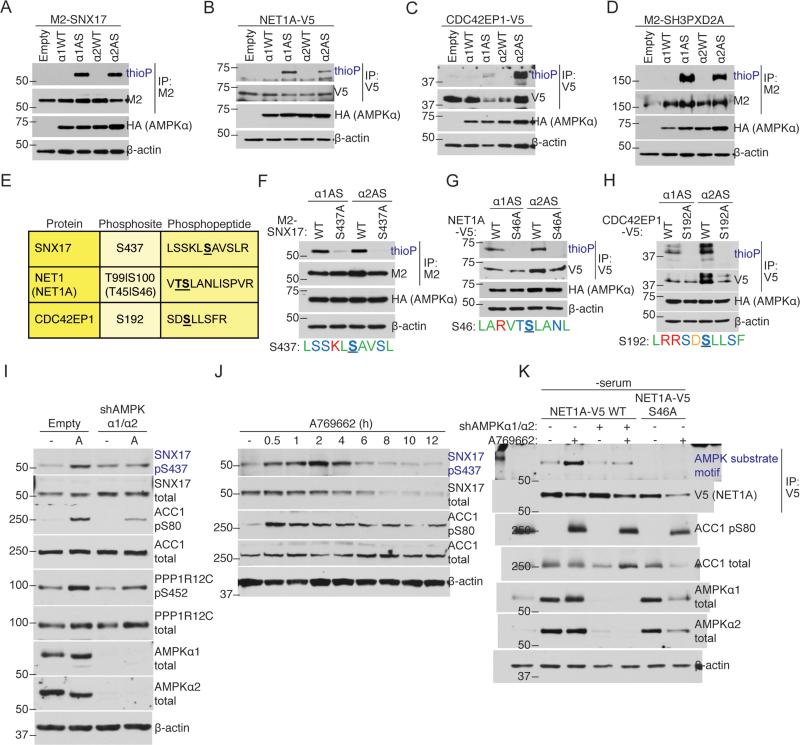
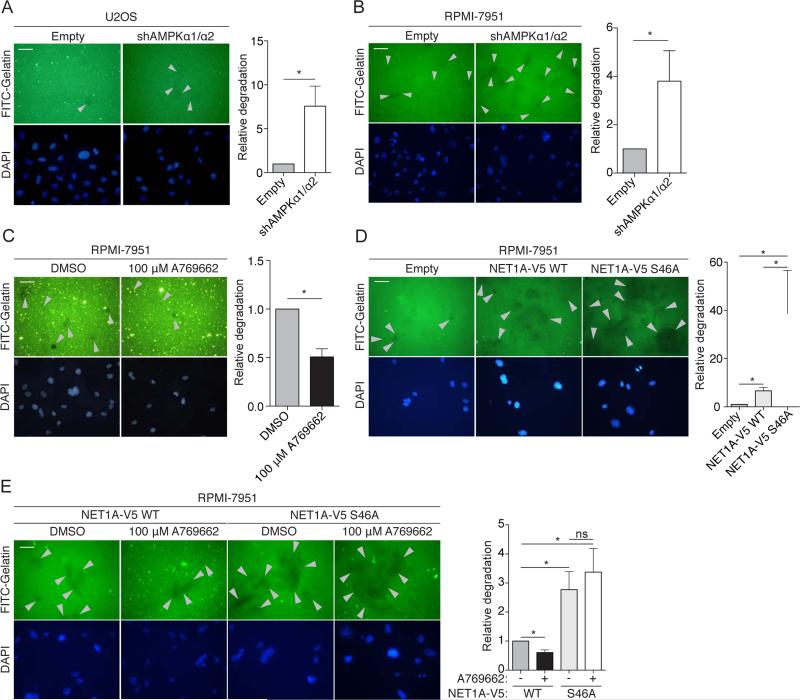
AMP-activated protein kinase (AMPK) is a central energy gauge that regulates metabolism and has been increasingly involved in non-metabolic processes and diseases. However, AMPK's direct substrates in non-metabolic contexts are largely unknown. To better understand the AMPK network, we use a chemical genetics screen coupled to a peptide capture approach in whole cells, resulting in identification of direct AMPK phosphorylation sites. Interestingly, the high-confidence AMPK substrates contain many proteins involved in cell motility, adhesion, and invasion. AMPK phosphorylation of the RHOA guanine nucleotide exchange factor NET1A inhibits extracellular matrix degradation, an early step in cell invasion. The identification of direct AMPK phosphorylation sites also facilitates large-scale prediction of AMPK substrates. We provide an AMPK motif matrix and a pipeline to predict additional AMPK substrates from quantitative phosphoproteomics datasets. As AMPK is emerging as a critical node in aging and pathological processes, our study identifies potential targets for therapeutic strategies.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures

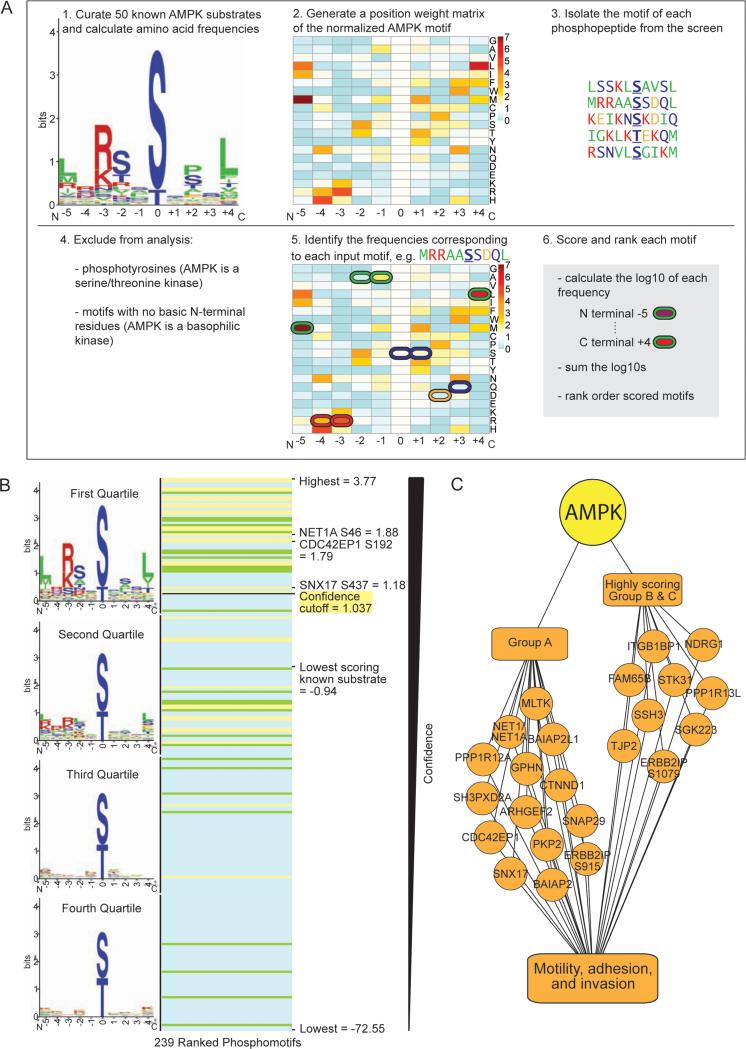
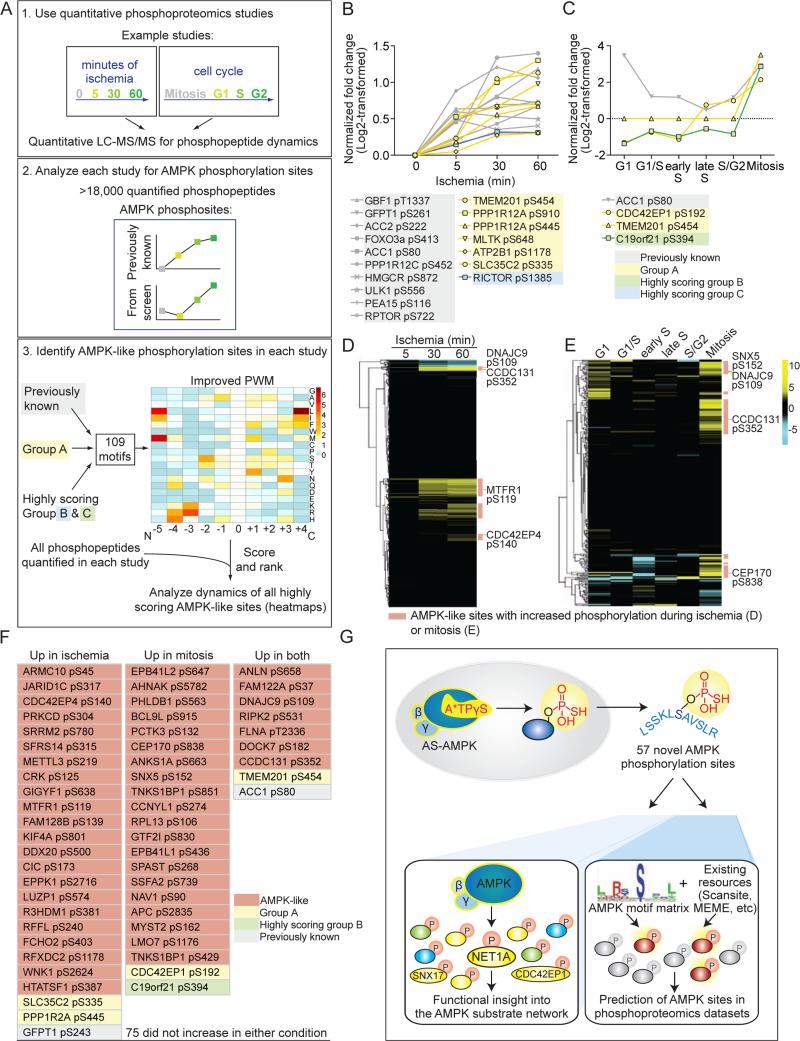
References
-
- Andersson U, Filipsson K, Abbott CR, Woods A, Smith K, Bloom SR, Carling D, Small CJ. AMP-activated protein kinase plays a role in the control of food intake. J Biol Chem. 2004;279:12005–12008. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- T32 CA009302/CA/NCI NIH HHS/United States
- P41 GM103481/GM/NIGMS NIH HHS/United States
- 8P41GM103481/GM/NIGMS NIH HHS/United States
- P41GM103481/GM/NIGMS NIH HHS/United States
- T32 CA09302/CA/NCI NIH HHS/United States
- R01 DK080425/DK/NIDDK NIH HHS/United States
- P01 CA120964/CA/NCI NIH HHS/United States
- R01 CA172229/CA/NCI NIH HHS/United States
- R01 AG031198/AG/NIA NIH HHS/United States
- S10 RR027425/RR/NCRR NIH HHS/United States
- S10RR027425/RR/NCRR NIH HHS/United States
- P01-AG032953/AG/NIA NIH HHS/United States
- P30 CA014195/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
