

CSNAP Is a Stoichiometric Subunit of the COP9 Signalosome
- PMID: 26456823
- PMCID: PMC5724754
- DOI: 10.1016/j.celrep.2015.09.021
CSNAP Is a Stoichiometric Subunit of the COP9 Signalosome
Abstract
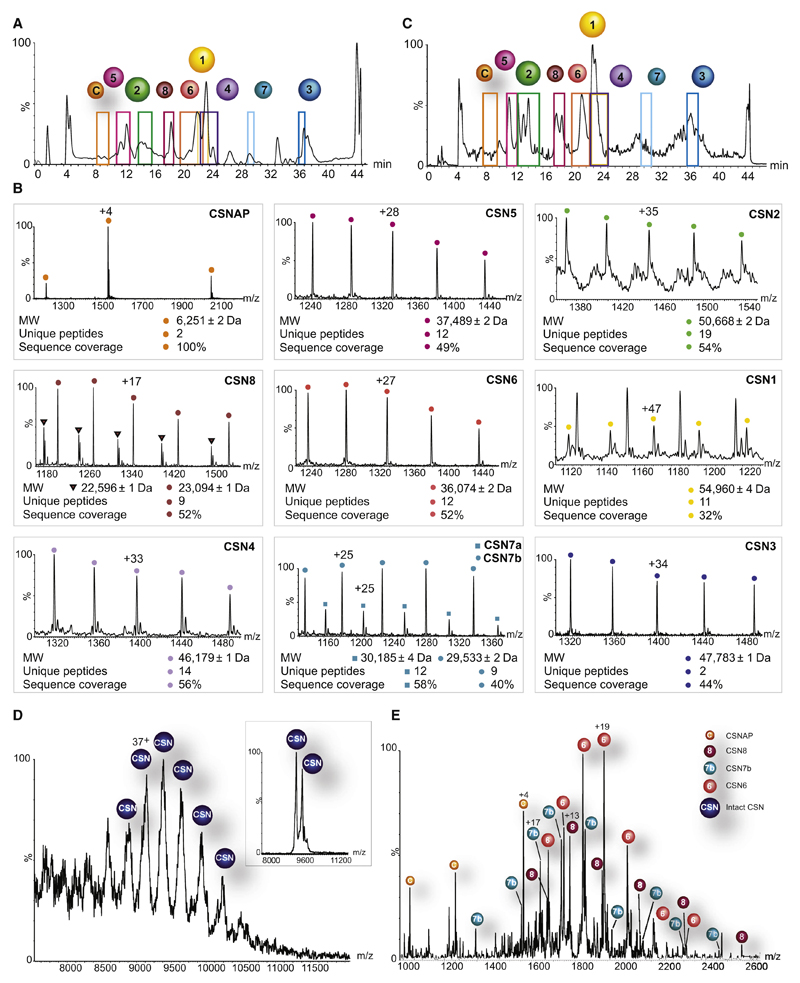
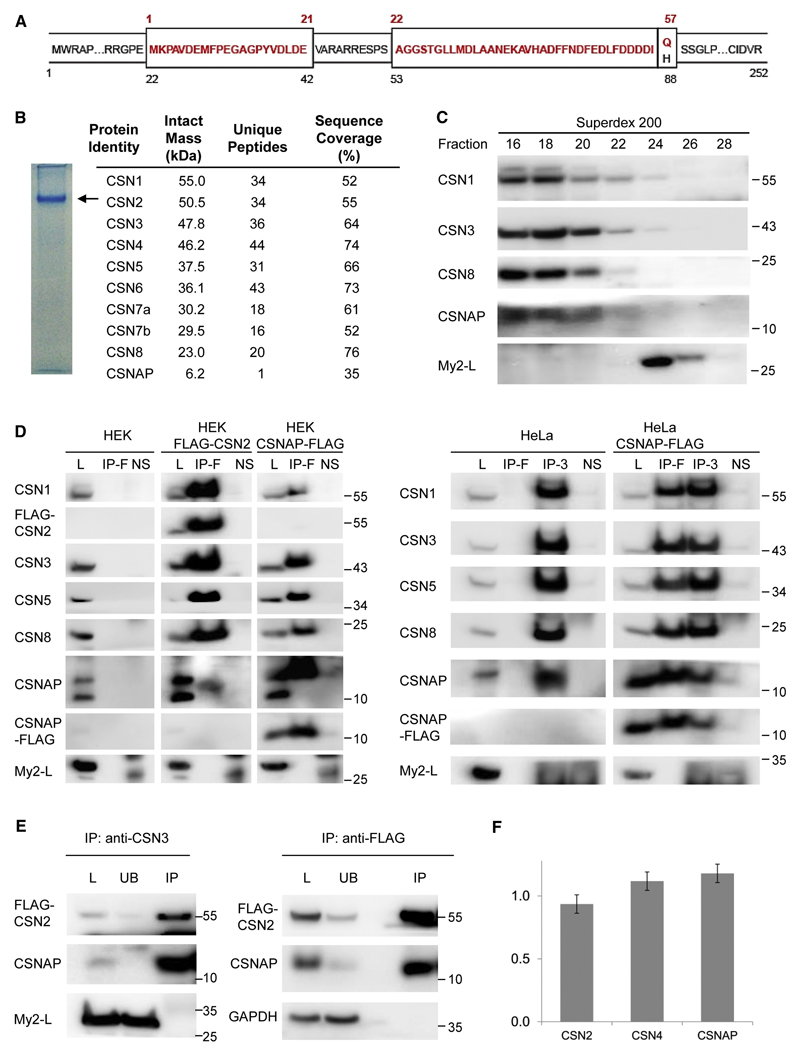
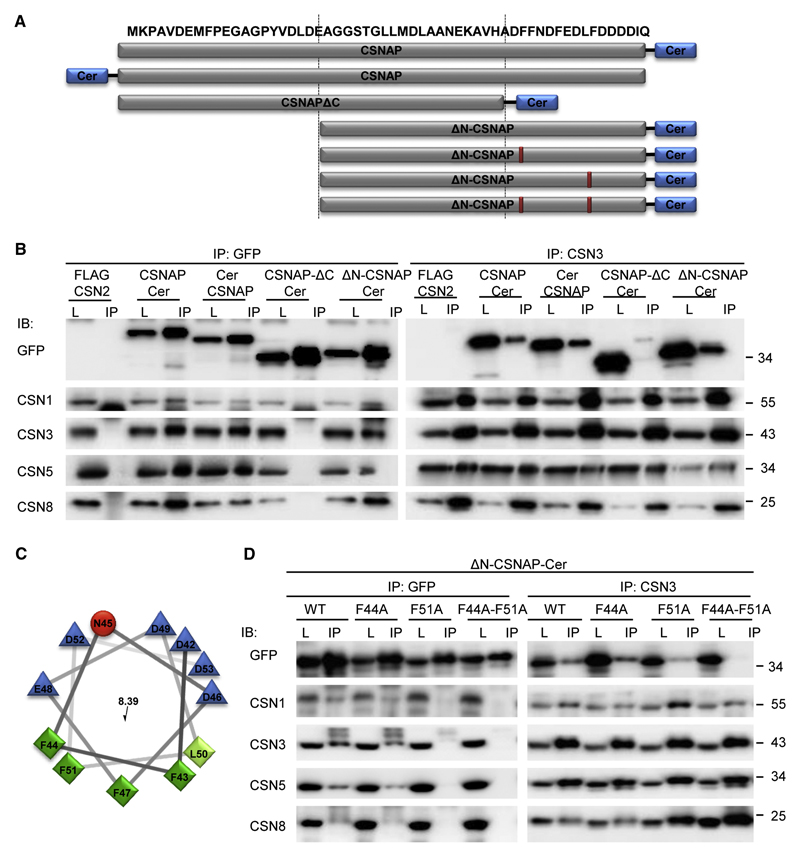
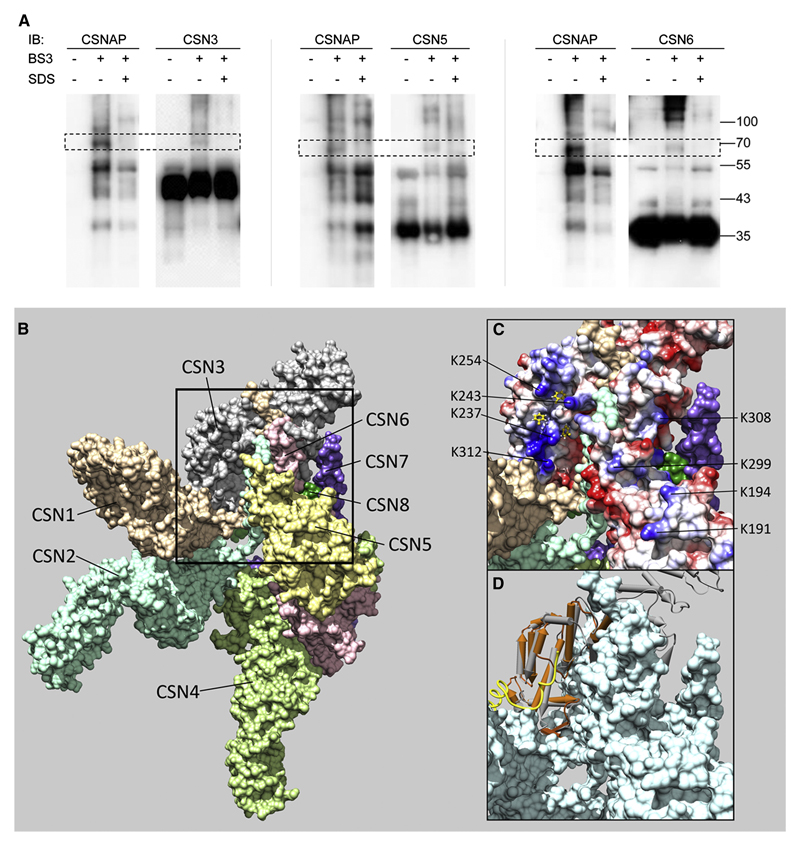
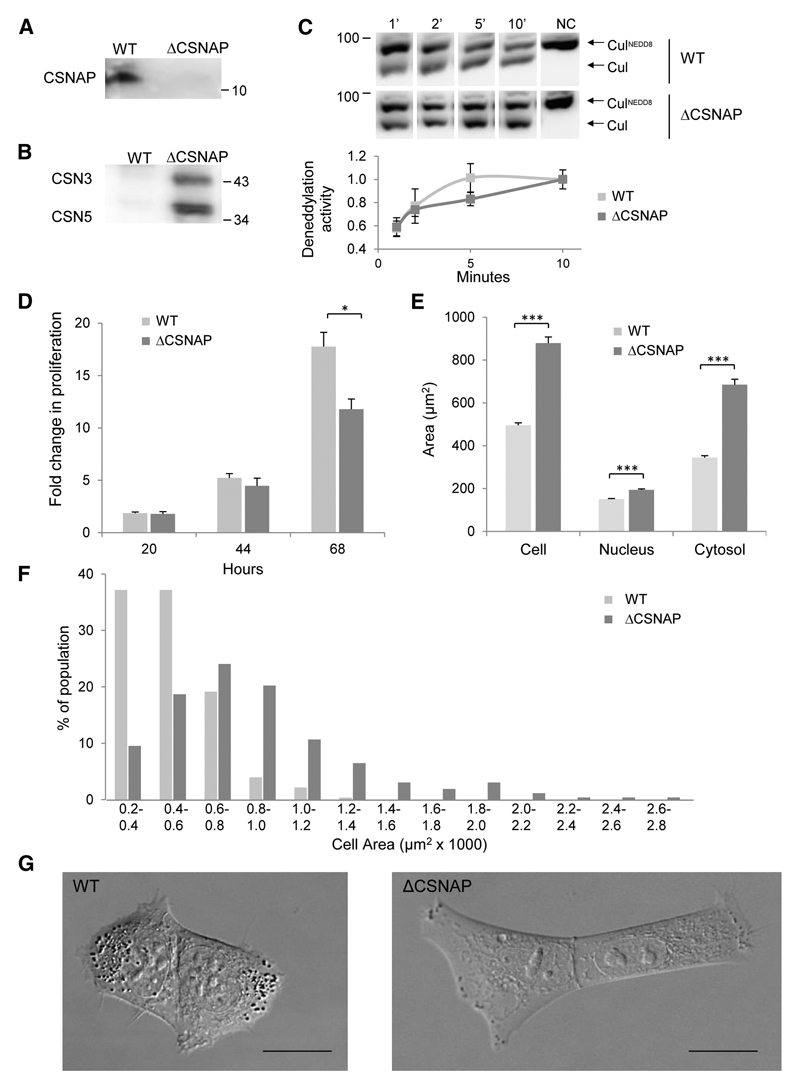
The highly conserved COP9 signalosome (CSN) complex is a key regulator of all cullin-RING-ubiquitin ligases (CRLs), the largest family of E3 ubiquitin ligases. Until now, it was accepted that the CSN is composed of eight canonical components. Here, we report the discovery of an additional integral and stoichiometric subunit that had thus far evaded detection, and we named it CSNAP (CSN acidic protein). We show that CSNAP binds CSN3, CSN5, and CSN6, and its incorporation into the CSN complex is mediated through the C-terminal region involving conserved aromatic residues. Moreover, depletion of this small protein leads to reduced proliferation and a flattened and enlarged morphology. Finally, on the basis of sequence and structural properties shared by both CSNAP and DSS1, a component of the related 19S lid proteasome complex, we propose that CSNAP, the ninth CSN subunit, is the missing paralogous subunit of DSS1.
Copyright © 2015 The Authors. Published by Elsevier Inc. All rights reserved.
Figures


Similar articles
-
Structural and biochemical characterization of the Cop9 signalosome CSN5/CSN6 heterodimer.PLoS One. 2014 Aug 21;9(8):e105688. doi: 10.1371/journal.pone.0105688. eCollection 2014. PLoS One. 2014. PMID: 25144743 Free PMC article.
-
Crystal structure of the human COP9 signalosome.Nature. 2014 Aug 14;512(7513):161-5. doi: 10.1038/nature13566. Epub 2014 Jul 16. Nature. 2014. PMID: 25043011
-
The minimal deneddylase core of the COP9 signalosome excludes the Csn6 MPN- domain.PLoS One. 2012;7(8):e43980. doi: 10.1371/journal.pone.0043980. Epub 2012 Aug 30. PLoS One. 2012. PMID: 22956996 Free PMC article.
-
Roles of COP9 signalosome in cancer.Cell Cycle. 2011 Sep 15;10(18):3057-66. doi: 10.4161/cc.10.18.17320. Epub 2011 Sep 15. Cell Cycle. 2011. PMID: 21876386 Free PMC article. Review.
-
The COP9 signalosome: A versatile regulatory hub of Cullin-RING ligases.Trends Biochem Sci. 2023 Jan;48(1):82-95. doi: 10.1016/j.tibs.2022.08.003. Epub 2022 Aug 27. Trends Biochem Sci. 2023. PMID: 36041947 Review.
Cited by
-
The disordered PCI-binding human proteins CSNAP and DSS1 have diverged in structure and function.Protein Sci. 2021 Oct;30(10):2069-2082. doi: 10.1002/pro.4159. Epub 2021 Jul 26. Protein Sci. 2021. PMID: 34272906 Free PMC article.
-
COP9 Signalosome CSN4 and CSN5 Subunits Are Involved in Jasmonate-Dependent Defense Against Root-Knot Nematode in Tomato.Front Plant Sci. 2019 Oct 9;10:1223. doi: 10.3389/fpls.2019.01223. eCollection 2019. Front Plant Sci. 2019. Retraction in: Front Plant Sci. 2025 Jul 30;16:1666310. doi: 10.3389/fpls.2025.1666310. PMID: 31649695 Free PMC article. Retracted.
-
Fungal COP9 signalosome assembly requires connection of two trimeric intermediates for integration of intrinsic deneddylase.Proc Natl Acad Sci U S A. 2023 Aug 29;120(35):e2305049120. doi: 10.1073/pnas.2305049120. Epub 2023 Aug 21. Proc Natl Acad Sci U S A. 2023. PMID: 37603767 Free PMC article.
-
Characterizing Endogenous Protein Complexes with Biological Mass Spectrometry.Chem Rev. 2022 Apr 27;122(8):7386-7414. doi: 10.1021/acs.chemrev.1c00217. Epub 2021 Aug 18. Chem Rev. 2022. PMID: 34406752 Free PMC article. Review.
-
Expanded Interactome of the Intrinsically Disordered Protein Dss1.Cell Rep. 2018 Oct 23;25(4):862-870. doi: 10.1016/j.celrep.2018.09.080. Cell Rep. 2018. PMID: 30355493 Free PMC article.
References
-
- Ben-Shimon A, Eisenstein M. Computational mapping of anchoring spots on protein surfaces. J Mol Biol. 2010;402:259–277. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
