

PAR1 signaling regulates the retention and recruitment of EPCR-expressing bone marrow hematopoietic stem cells
- PMID: 26457757
- PMCID: PMC4776769
- DOI: 10.1038/nm.3960
PAR1 signaling regulates the retention and recruitment of EPCR-expressing bone marrow hematopoietic stem cells
Erratum in
-
Corrigendum: PAR1 signaling regulates the retention and recruitment of EPCR-expressing bone marrow hematopoietic stem cells.Nat Med. 2016 Apr;22(4):446. doi: 10.1038/nm0416-446b. Nat Med. 2016. PMID: 27050594 No abstract available.
Abstract
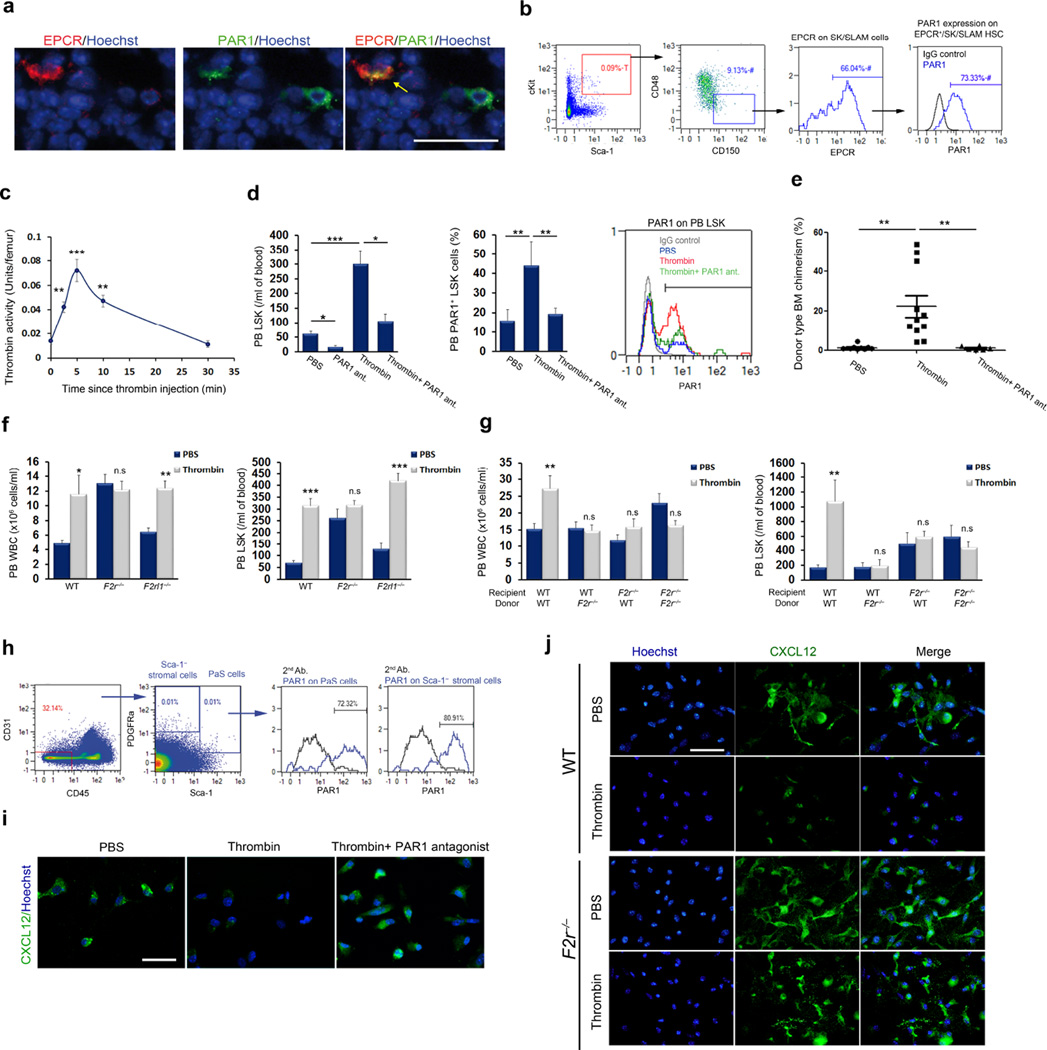
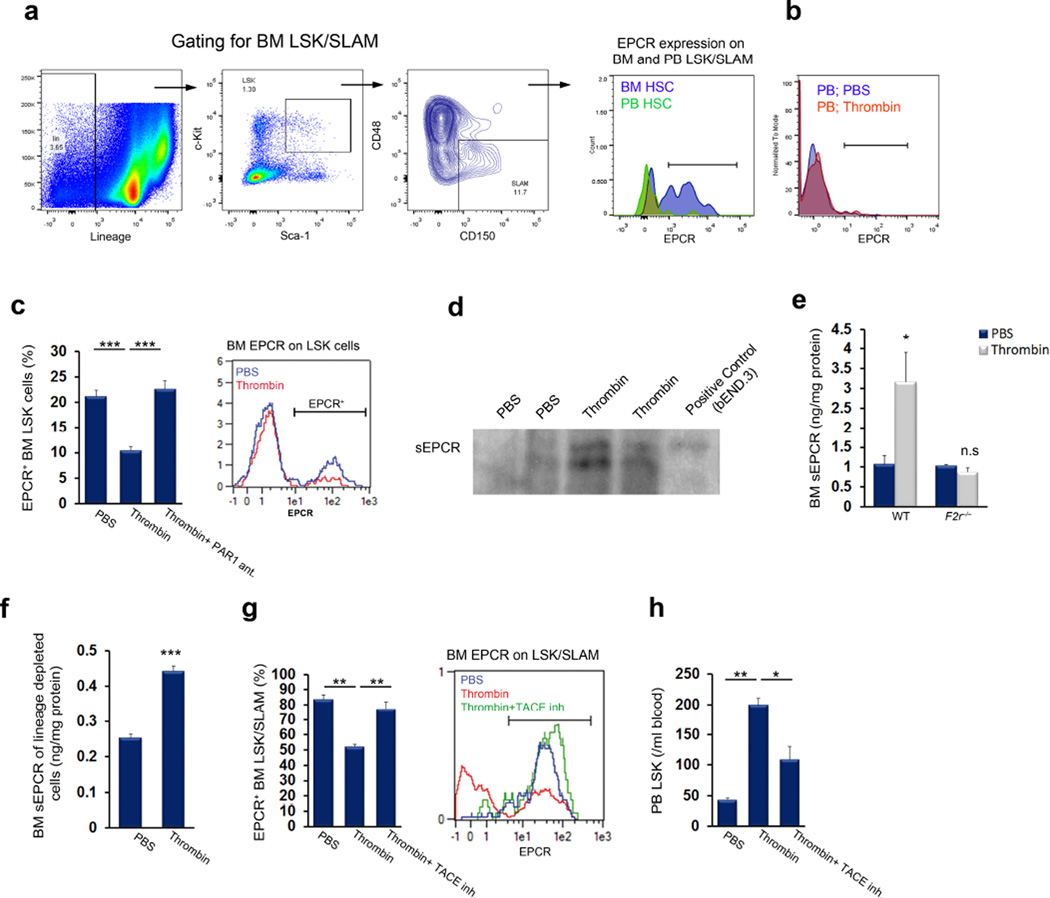
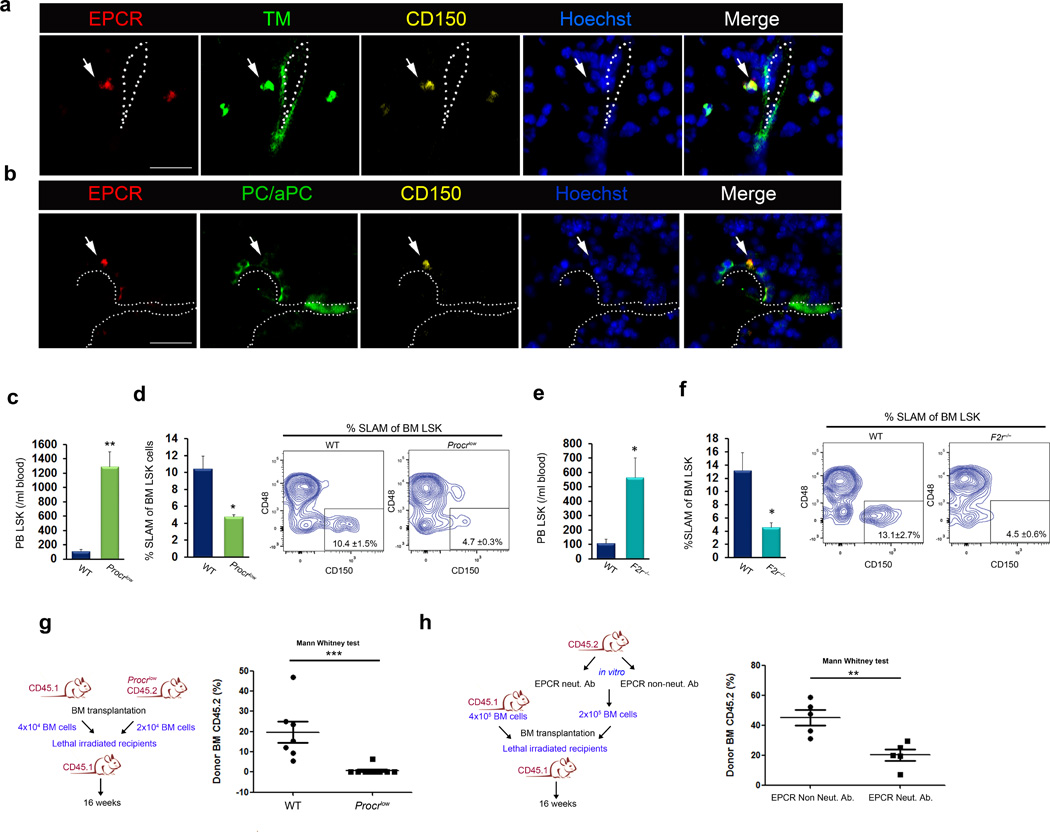
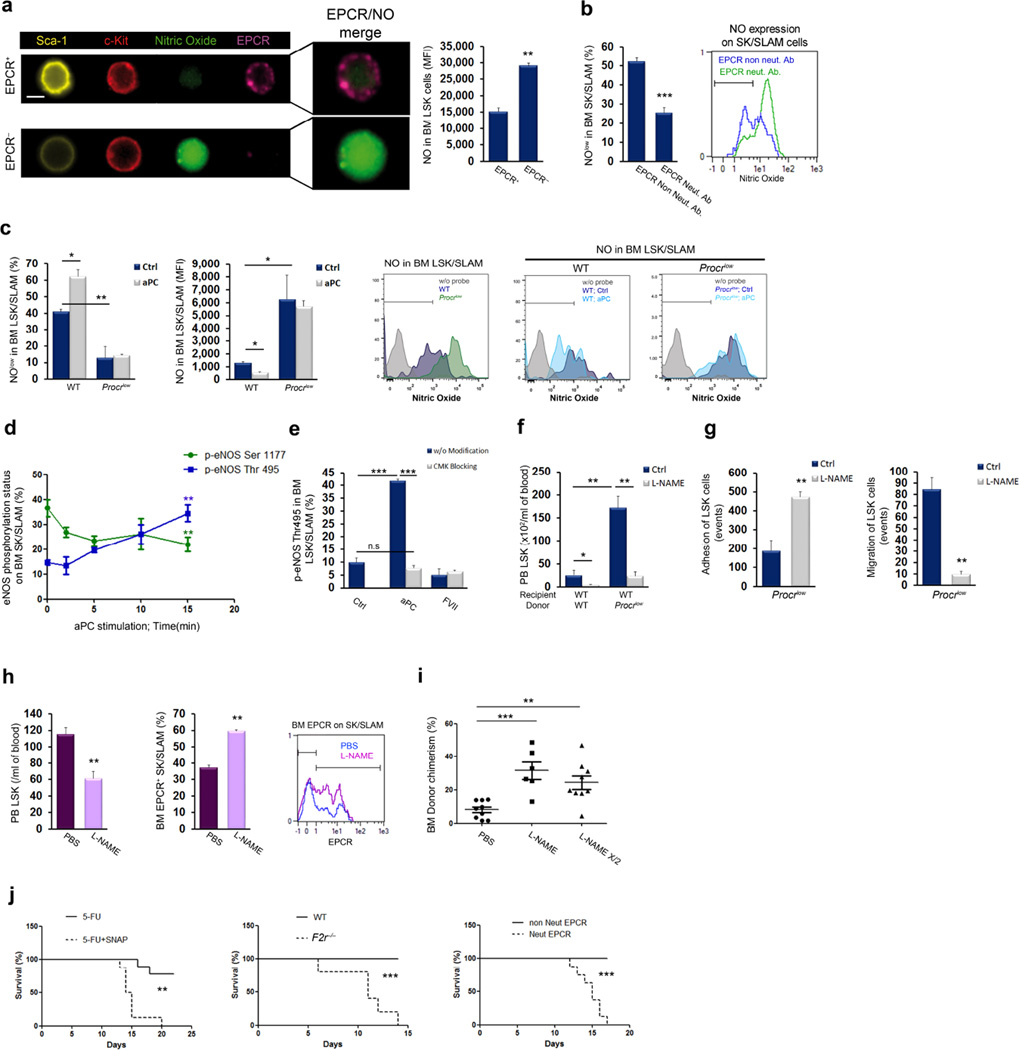
Retention of long-term repopulating hematopoietic stem cells (LT-HSCs) in the bone marrow is essential for hematopoiesis and for protection from myelotoxic injury. We report that signaling cascades that are traditionally viewed as coagulation related also control retention of endothelial protein C receptor-positive (EPCR(+)) LT-HSCs in the bone marrow and their recruitment to the blood via two pathways mediated by protease activated receptor 1 (PAR1). Thrombin-PAR1 signaling induces nitric oxide (NO) production, leading to EPCR shedding mediated by tumor necrosis factor-α-converting enzyme (TACE), enhanced CXCL12-CXCR4-induced motility and rapid stem and progenitor cell mobilization. Conversely, bone marrow blood vessels provide a microenvironment enriched with activated protein C (aPC) that retains EPCR(+) LT-HSCs by limiting NO generation, reducing Cdc42 activity and enhancing integrin VLA4 affinity and adhesion. Inhibition of NO production by aPC-EPCR-PAR1 signaling reduces progenitor cell egress from the bone marrow, increases retention of bone marrow NO(low) EPCR(+) LT-HSCs and protects mice from chemotherapy-induced hematological failure and death. Our study reveals new roles for PAR1 and EPCR in controlling NO production to balance maintenance and recruitment of bone marrow EPCR(+) LT-HSCs, with potential clinical relevance for stem cell transplantation.
Figures


References
-
- Spiegel A, Kalinkovich A, Shivtiel S, Kollet O, Lapidot T. Stem cell regulation via dynamic interactions of the nervous and immune systems with the microenvironment. Cell Stem Cell. 2008;3:484–492. - PubMed
-
- Sugiyama T, Kohara H, Noda M, Nagasawa T. Maintenance of the hematopoietic stem cell pool by CXCL12-CXCR4 chemokine signaling in bone marrow stromal cell niches. Immunity. 2006;25:977–988. - PubMed
-
- Schajnovitz A, et al. CXCL12 secretion by bone marrow stromal cells is dependent on cell contact and mediated by connexin-43 and connexin-45 gap junctions. Nat Immunol. 2011;12:391–398. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
