

Gating machinery of InsP3R channels revealed by electron cryomicroscopy
- PMID: 26458101
- PMCID: PMC4804758
- DOI: 10.1038/nature15249
Gating machinery of InsP3R channels revealed by electron cryomicroscopy
Abstract
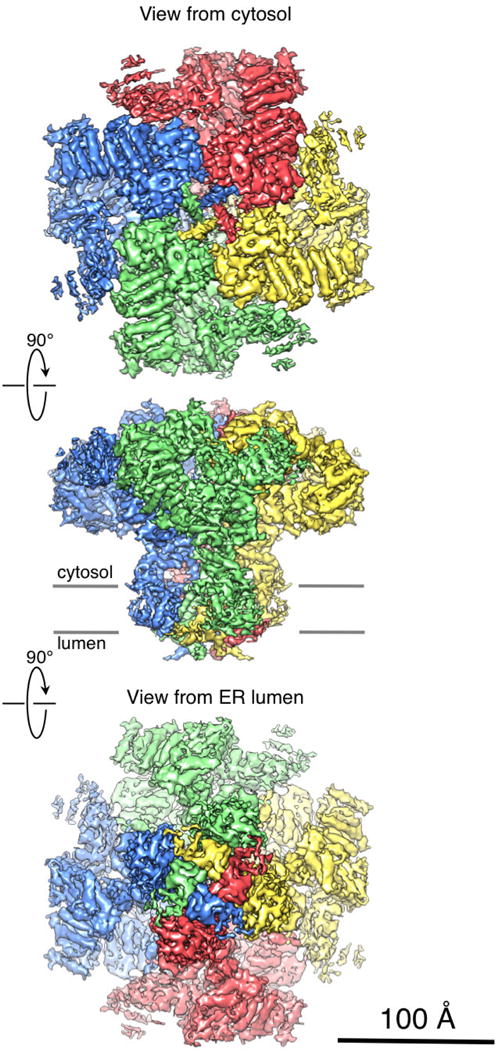
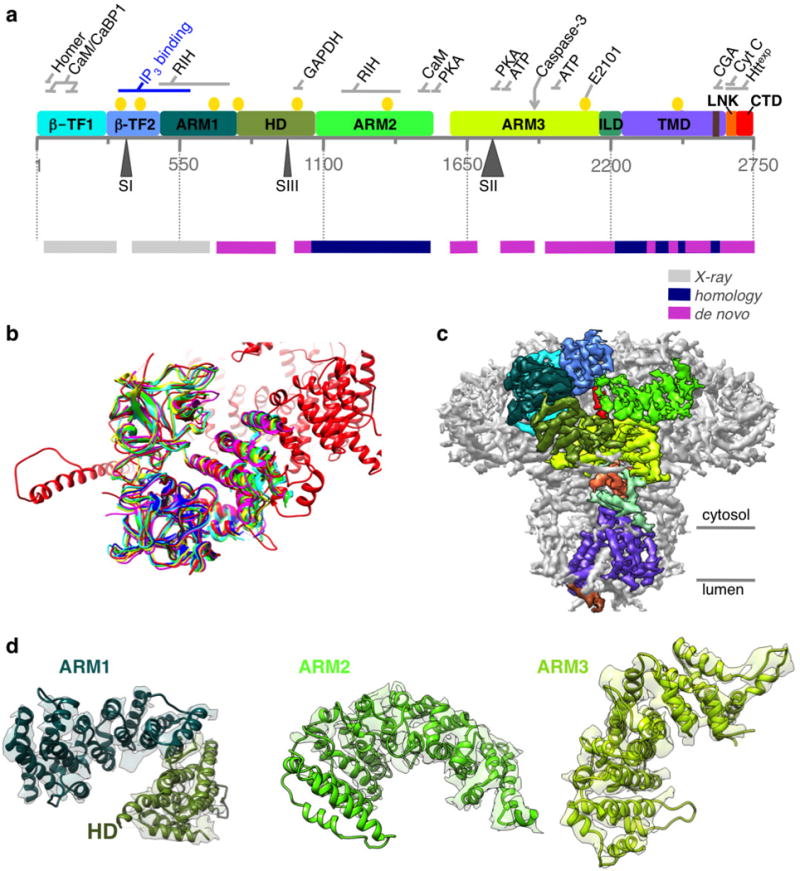
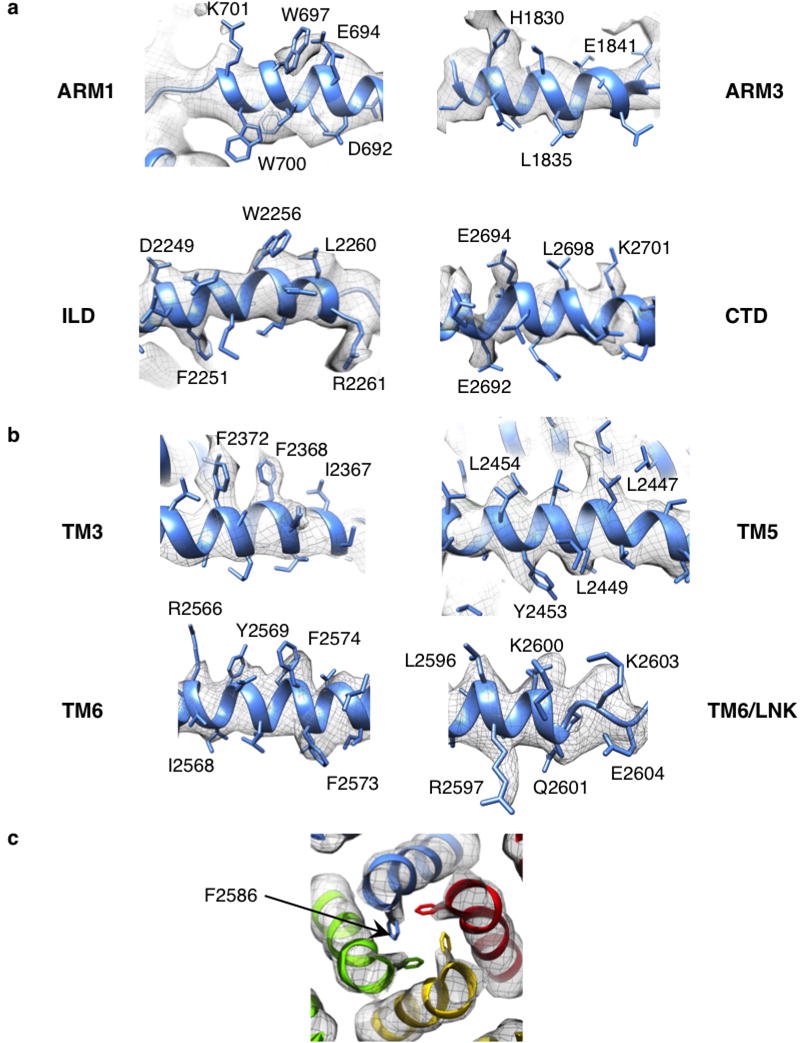
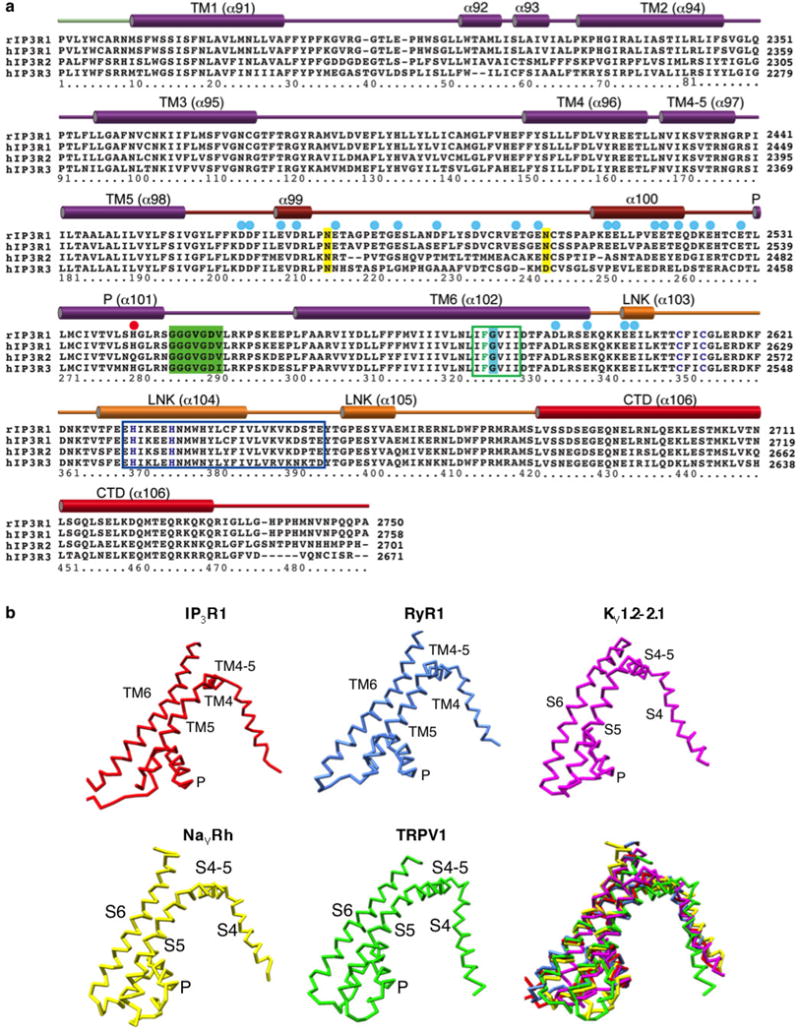
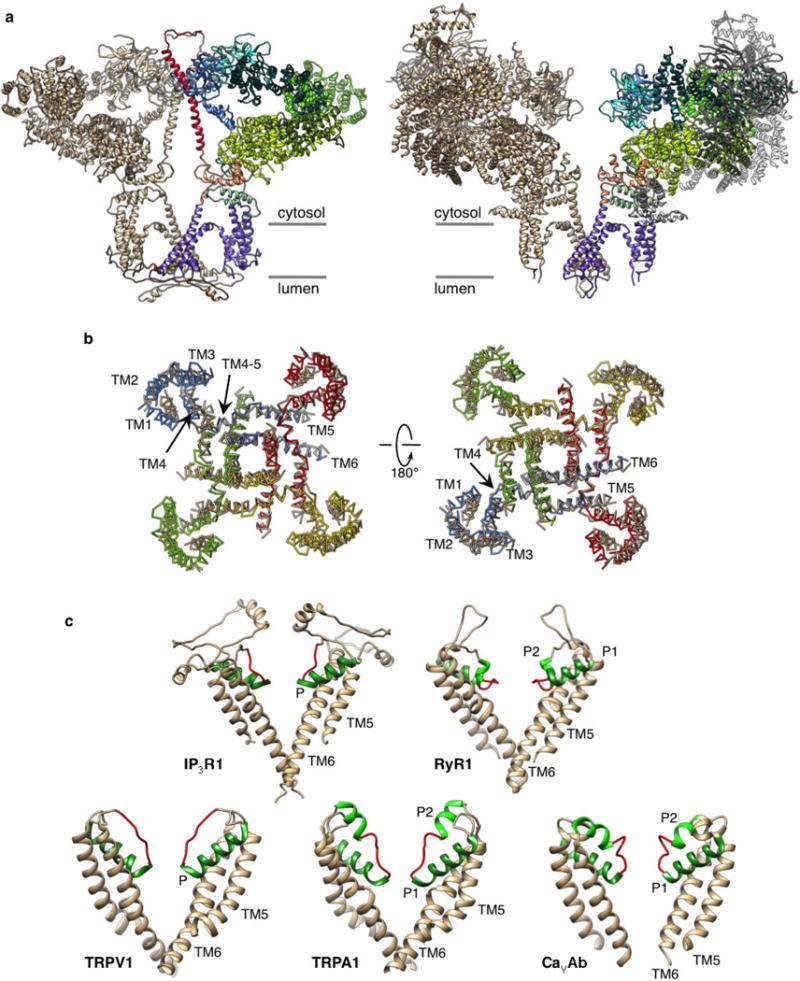
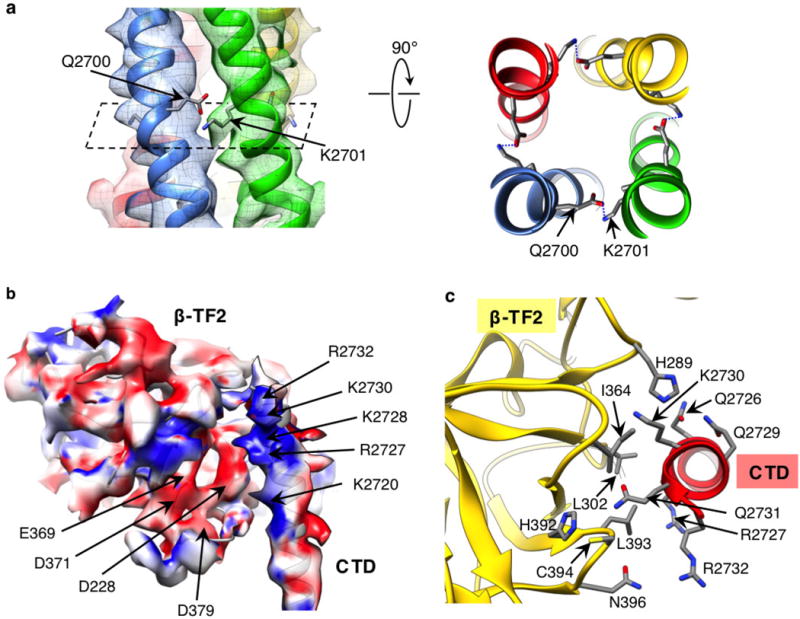
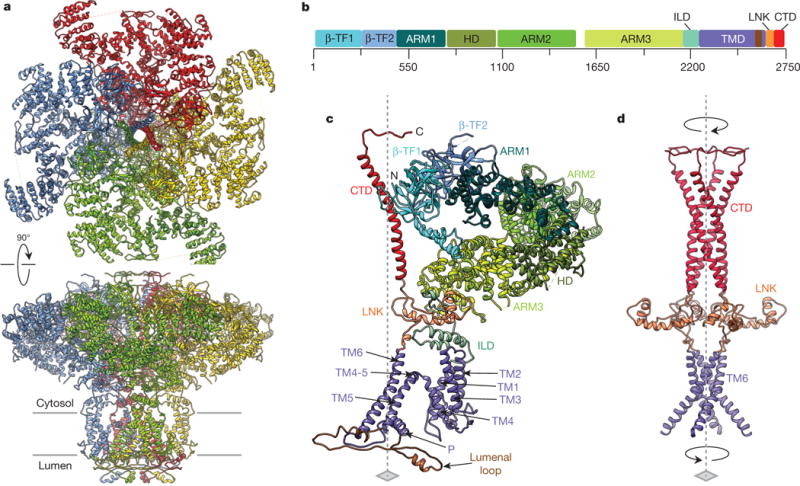
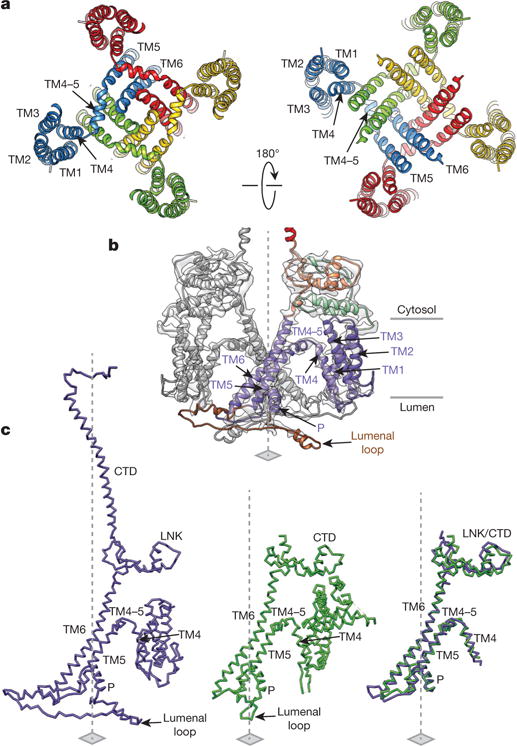
Inositol-1,4,5-trisphosphate receptors (InsP3Rs) are ubiquitous ion channels responsible for cytosolic Ca(2+) signalling and essential for a broad array of cellular processes ranging from contraction to secretion, and from proliferation to cell death. Despite decades of research on InsP3Rs, a mechanistic understanding of their structure-function relationship is lacking. Here we present the first, to our knowledge, near-atomic (4.7 Å) resolution electron cryomicroscopy structure of the tetrameric mammalian type 1 InsP3R channel in its apo-state. At this resolution, we are able to trace unambiguously ∼85% of the protein backbone, allowing us to identify the structural elements involved in gating and modulation of this 1.3-megadalton channel. Although the central Ca(2+)-conduction pathway is similar to other ion channels, including the closely related ryanodine receptor, the cytosolic carboxy termini are uniquely arranged in a left-handed α-helical bundle, directly interacting with the amino-terminal domains of adjacent subunits. This configuration suggests a molecular mechanism for allosteric regulation of channel gating by intracellular signals.
Conflict of interest statement
The authors declare no competing financial interests. Readers are welcome to comment on the online version of the paper.
Figures





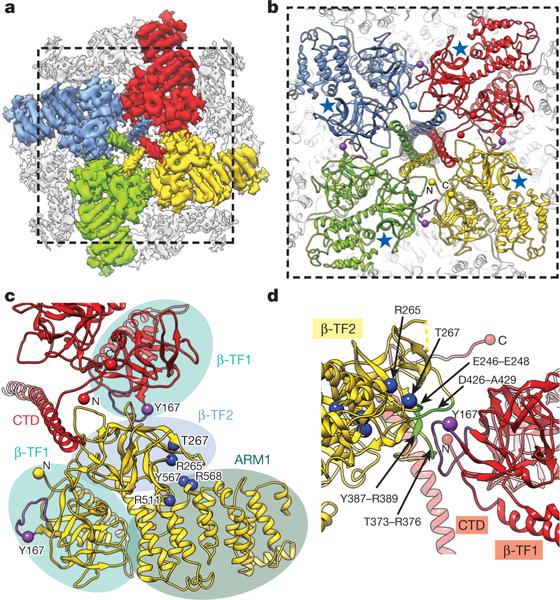
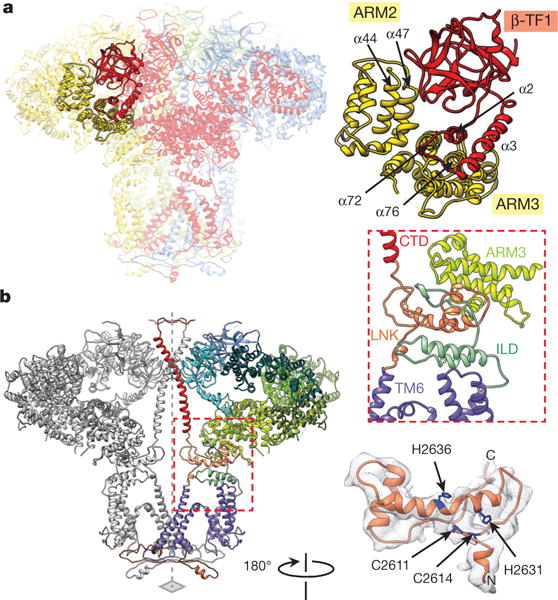
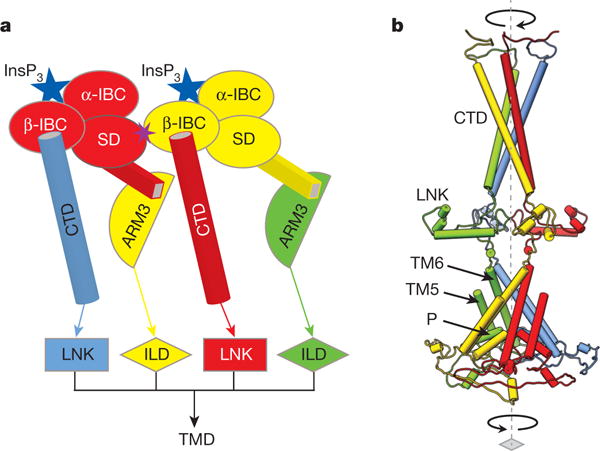
References
-
- Serysheva II, Ludtke SJ. 3D structure of IP3 receptor. Curr Topics Memb. 2010;66C:171–189. - PubMed
-
- Bosanac I, et al. Structure of the inositol 1,4,5-trisphosphate receptor binding core in complex with its ligand. Nature. 2002;420:696–700. - PubMed
-
- Bosanac I, et al. Crystal structure of the ligand binding suppressor domain of type 1 inositol 1,4,5-trisphosphate receptor. Mol Cell. 2005;17:193–203. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
- R01 GM080139/GM/NIGMS NIH HHS/United States
- R01GM080139/GM/NIGMS NIH HHS/United States
- R21 AR063255/AR/NIAMS NIH HHS/United States
- R01GM072804/GM/NIGMS NIH HHS/United States
- P41GM103832/GM/NIGMS NIH HHS/United States
- P41 GM103832/GM/NIGMS NIH HHS/United States
- R01 GM079429/GM/NIGMS NIH HHS/United States
- R21 GM100229/GM/NIGMS NIH HHS/United States
- R01 GM072804/GM/NIGMS NIH HHS/United States
- S10OD016279/OD/NIH HHS/United States
- R21GM100229/GM/NIGMS NIH HHS/United States
- R01GM079429/GM/NIGMS NIH HHS/United States
- S10 OD016279/OD/NIH HHS/United States
- R21AR063255/AR/NIAMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
