

Unique and redundant functions of NKp46+ ILC3s in models of intestinal inflammation
- PMID: 26458769
- PMCID: PMC4612098
- DOI: 10.1084/jem.20151403
Unique and redundant functions of NKp46+ ILC3s in models of intestinal inflammation
Abstract
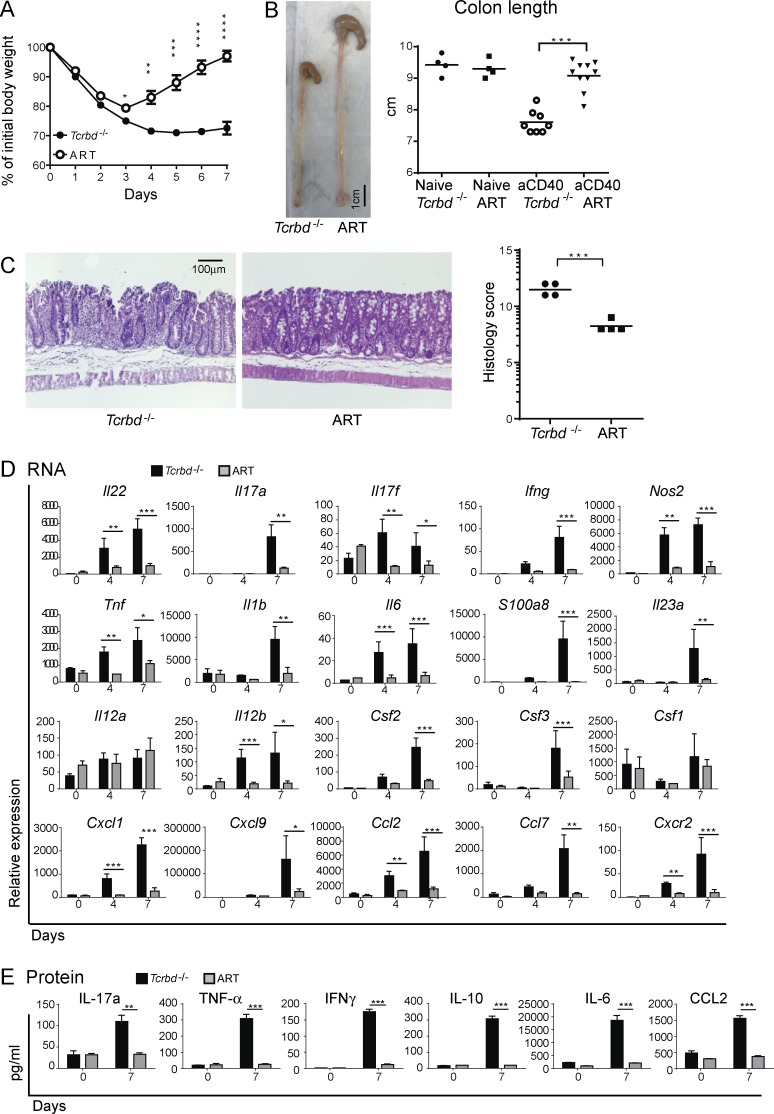
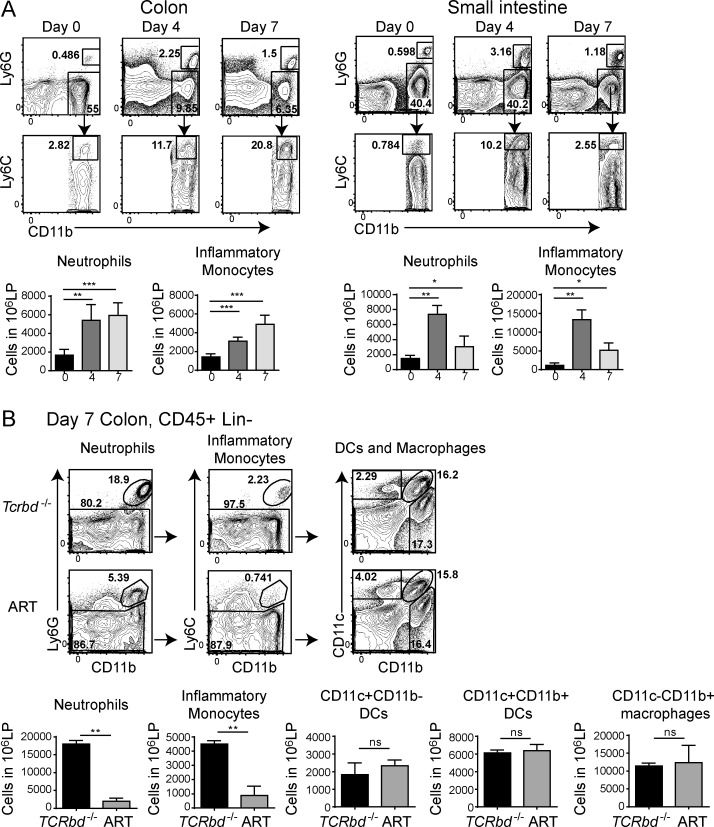
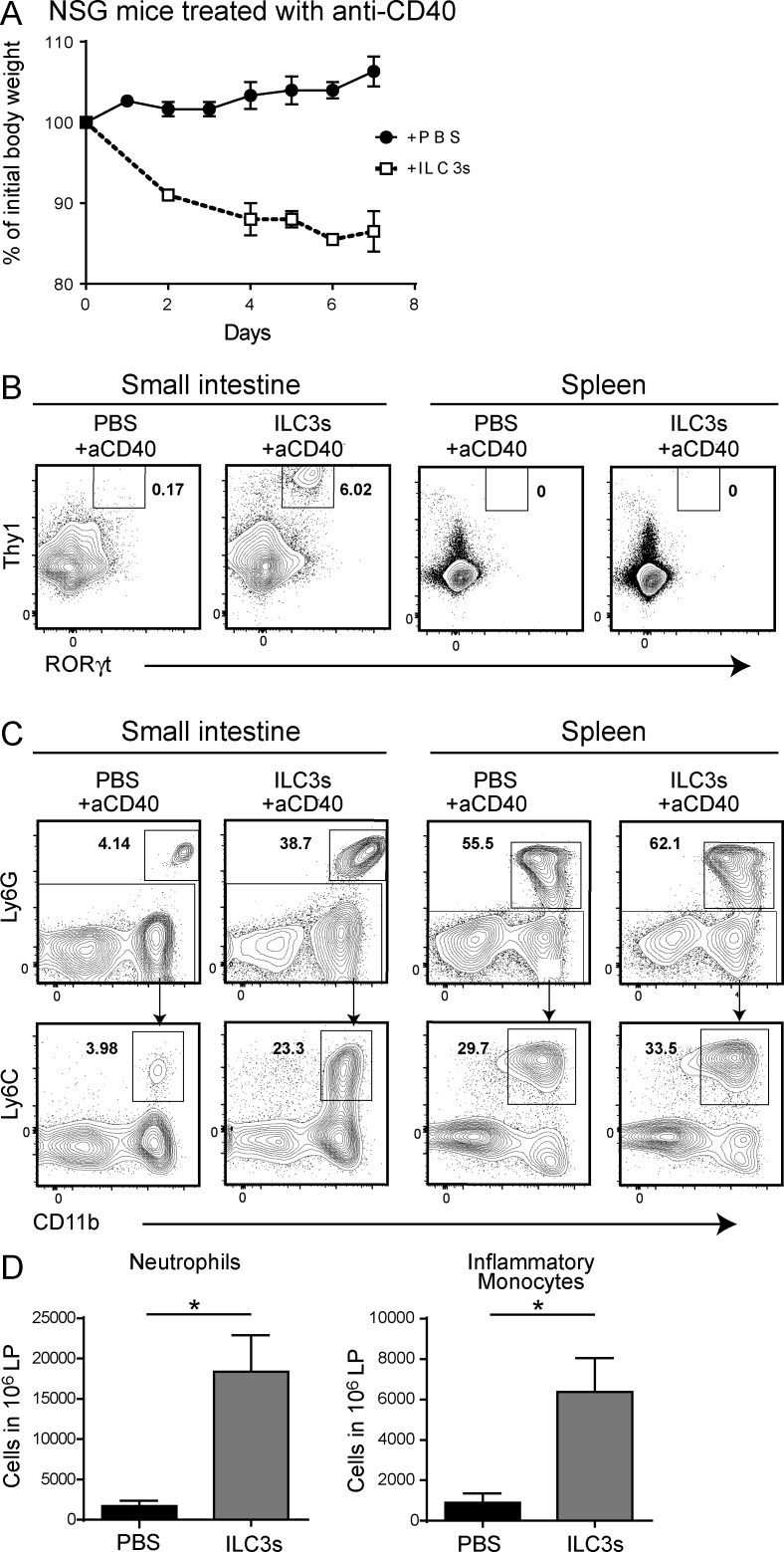
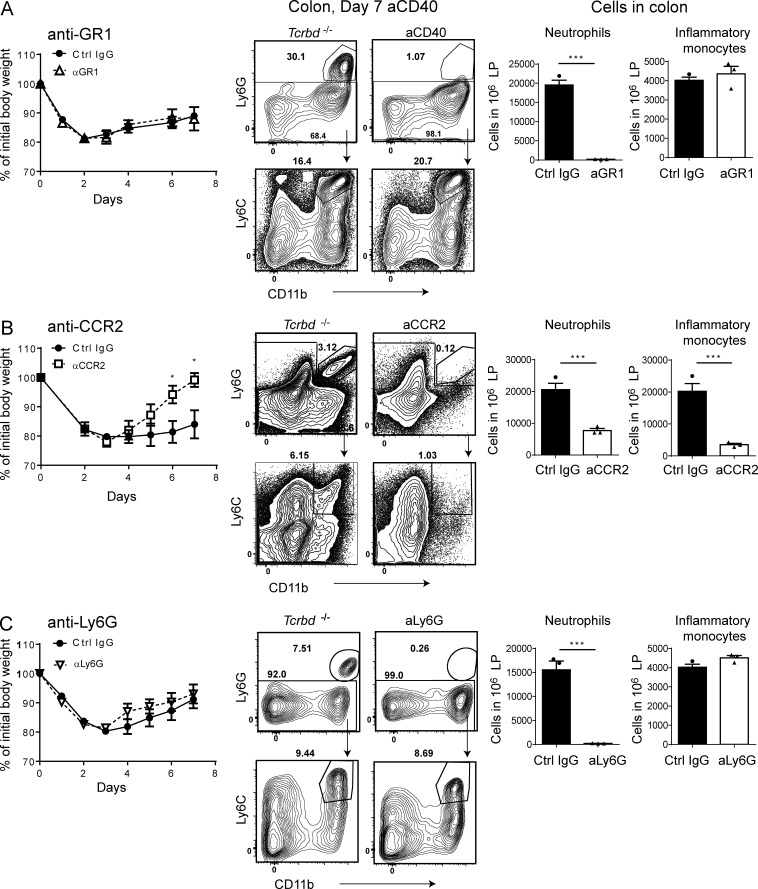
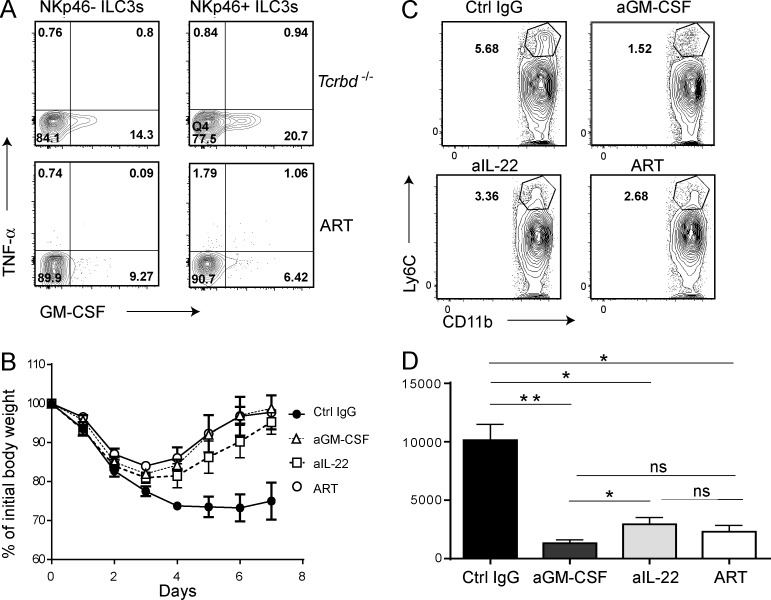
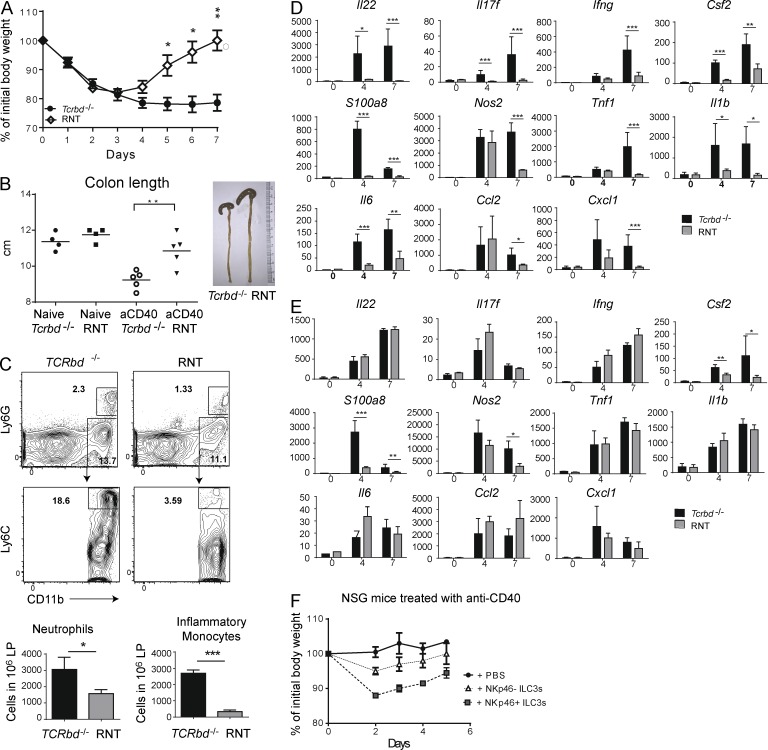
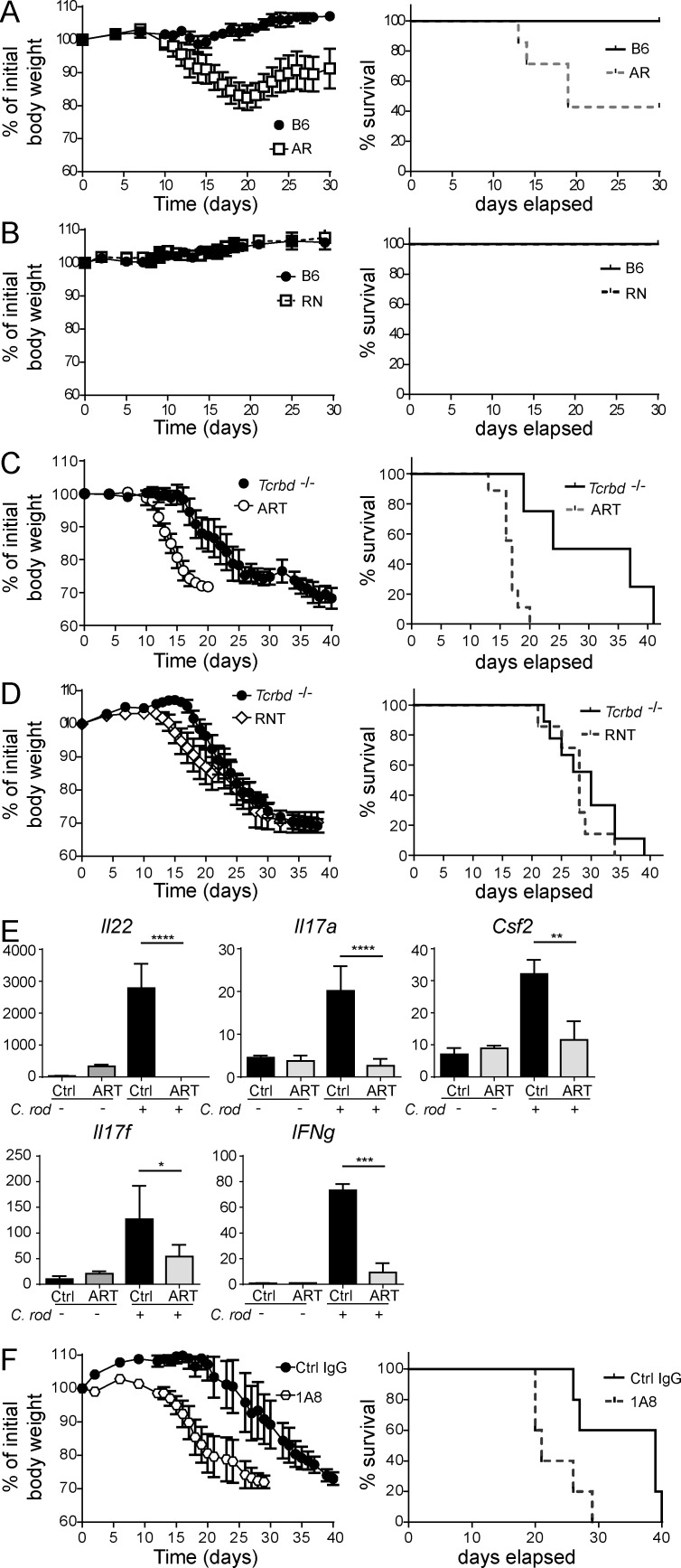
Group 3 ILCs (ILC3s) are innate sources of IL-22 and IL-17 and include lymphoid tissue-inducer (LTi)-like and NKp46(+) subsets. Both depend on RORγt and aryl hydrocarbon receptor, but NKp46(+)ILC3s also require Notch and T-bet for their development and are transcriptionally distinct. The extent to which these subsets have unique functions, especially in the context of T cell- and B cell-sufficient mice, remains largely unclear. To investigate the specific function of NKp46(+)ILC3s among other ILC3 subsets and T cells, we generated mice selectively lacking NKp46(+)ILC3s or all ILC3s and crossed them to T cell-deficient mice, thus maintaining B cells in all mice. In mice lacking T cells, NKp46(+)ILC3s were sufficient to promote inflammatory monocyte accumulation in the anti-CD40 innate colitis model through marked production of GM-CSF. In T cell-competent mice, lack of NKp46(+)ILCs had no impact on control of intestinal C. rodentium infection, whereas lack of all ILC3s partially impaired bacterial control. Thus, NKp46(+)ILC3s have a unique capacity to promote inflammation through GM-CSF-induced accumulation of inflammatory monocytes, but are superseded by LTi-like ILC3s and T cells in controlling intestinal bacterial infection.
© 2015 Song et al.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
