

The Solution Structure of the Lantibiotic Immunity Protein NisI and Its Interactions with Nisin
- PMID: 26459561
- PMCID: PMC4661402
- DOI: 10.1074/jbc.M115.679969
The Solution Structure of the Lantibiotic Immunity Protein NisI and Its Interactions with Nisin
Abstract
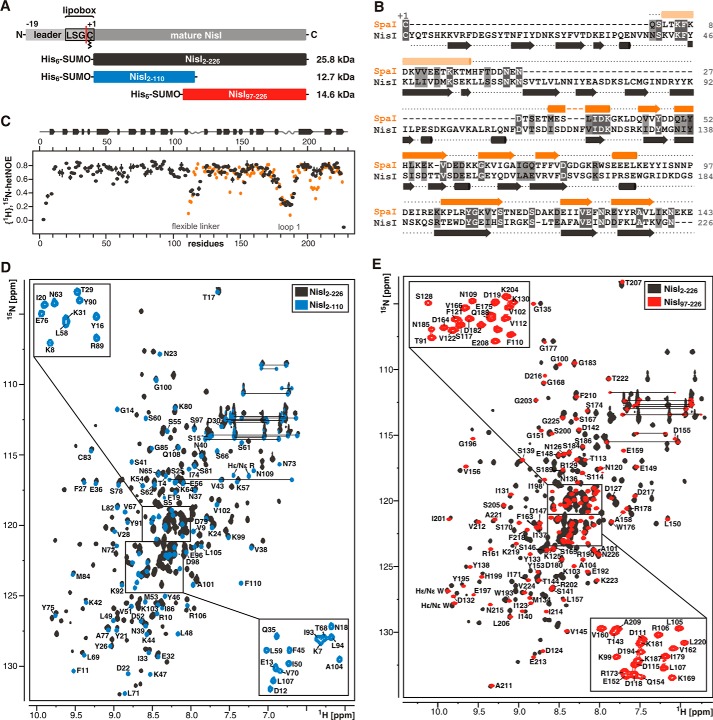
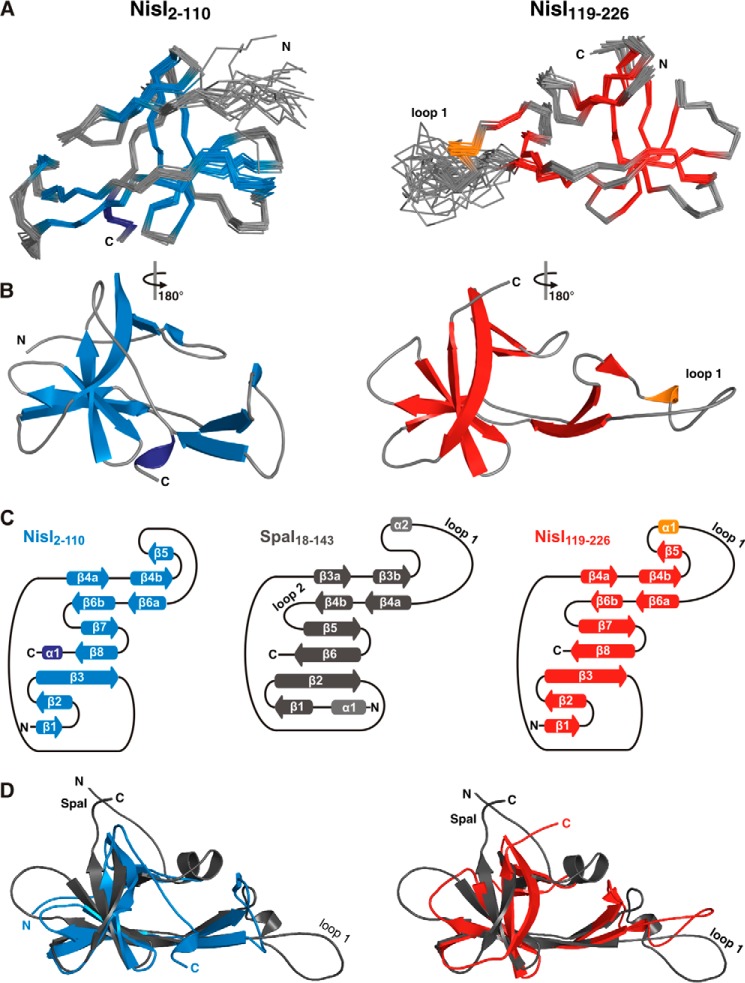
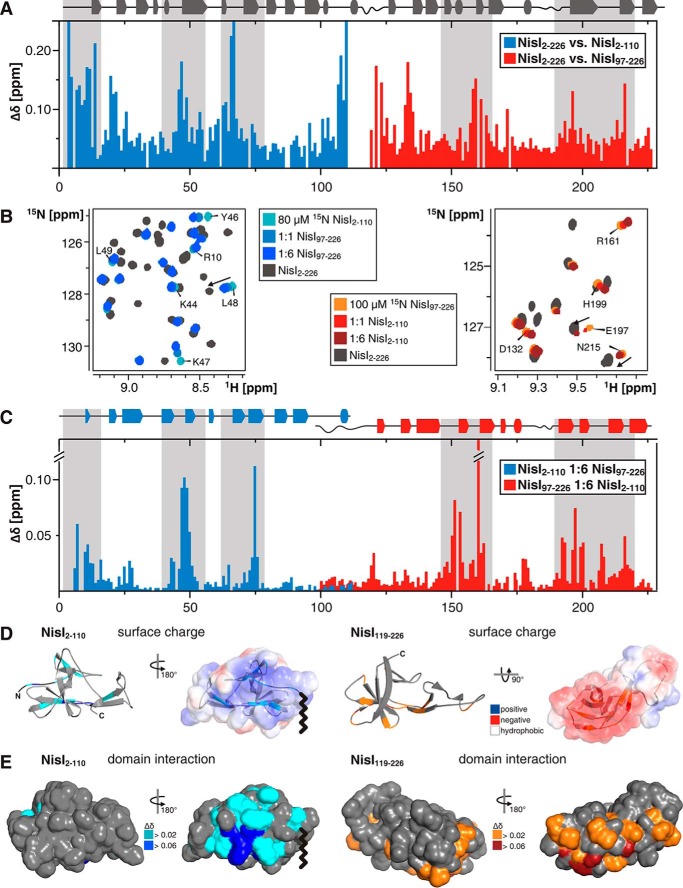
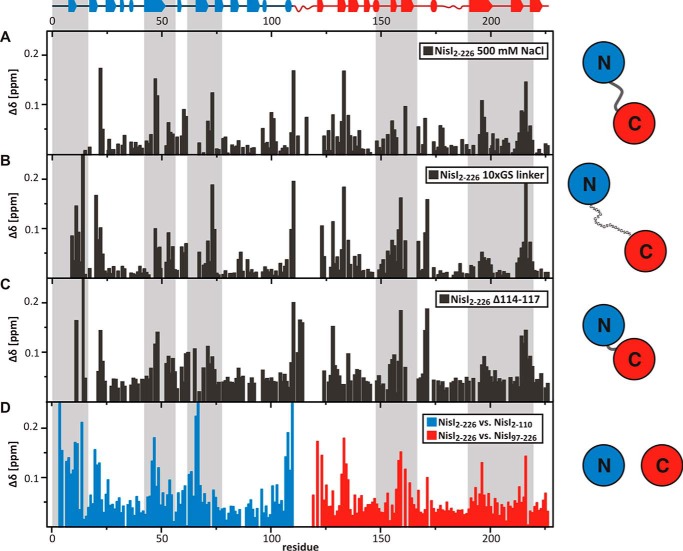
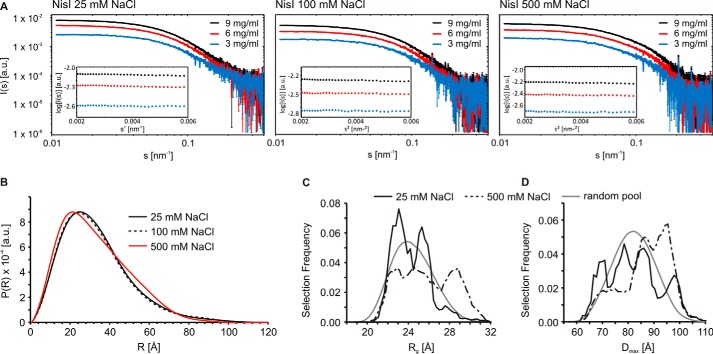
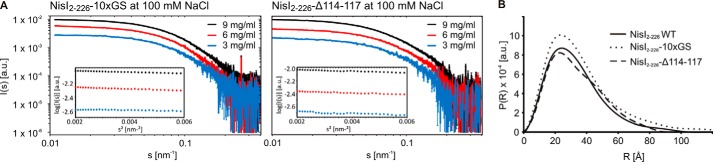
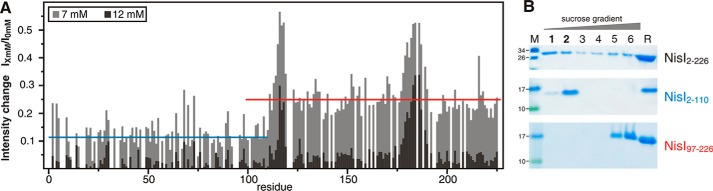
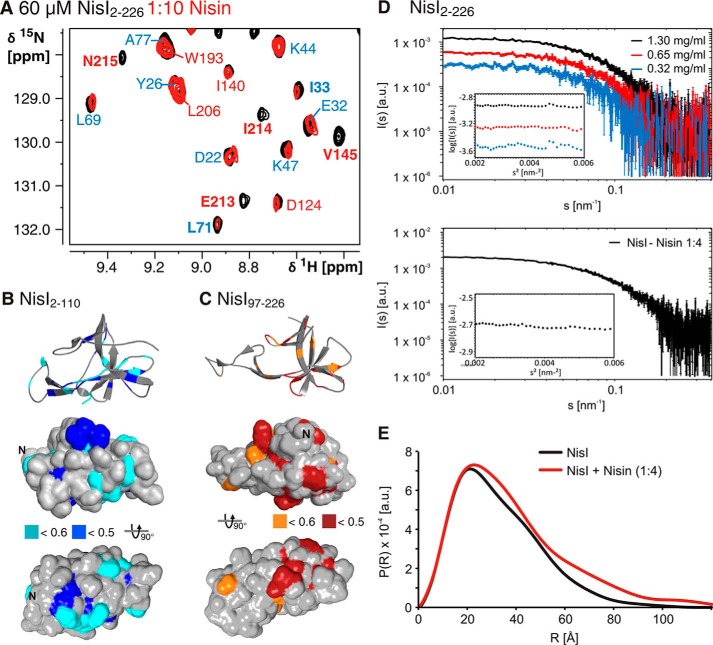
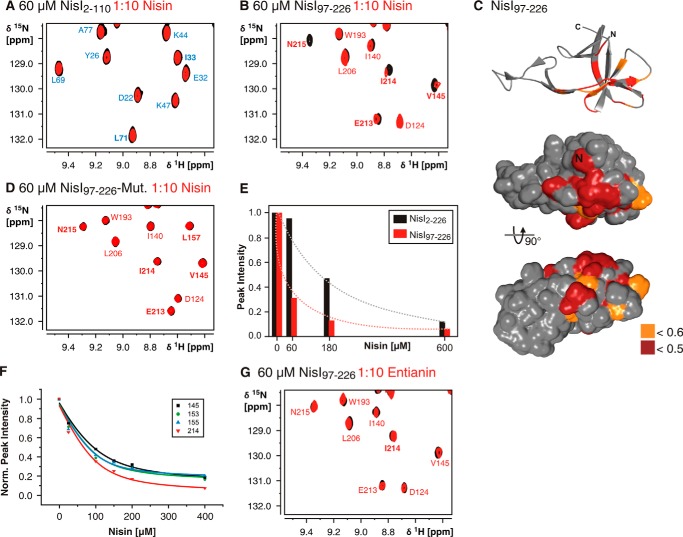
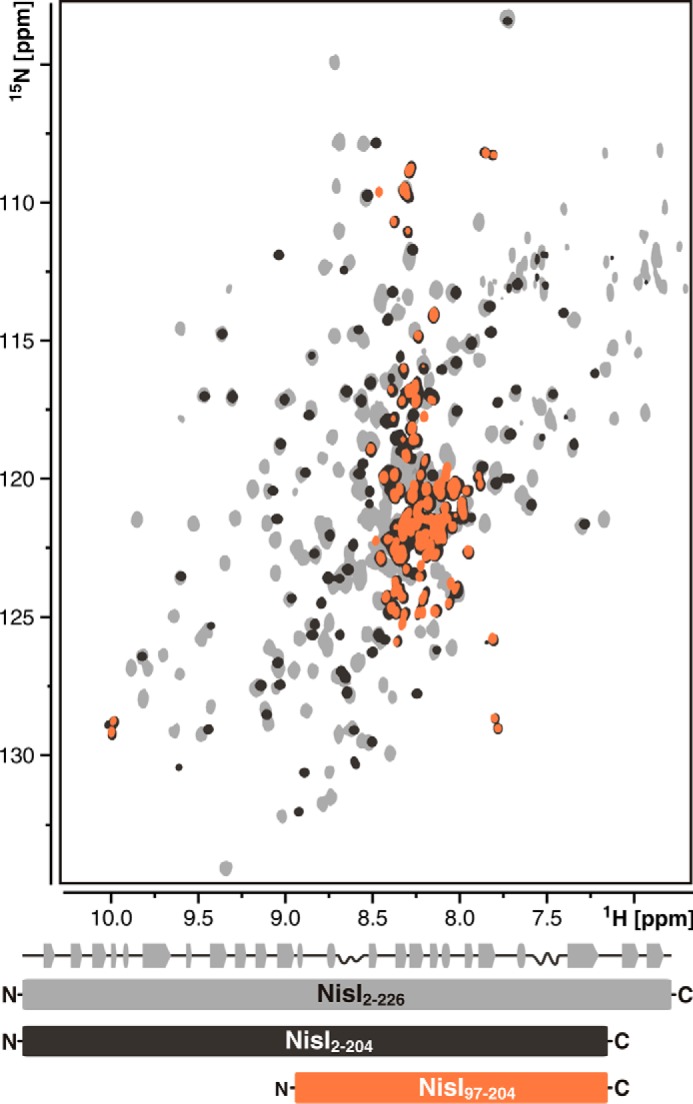
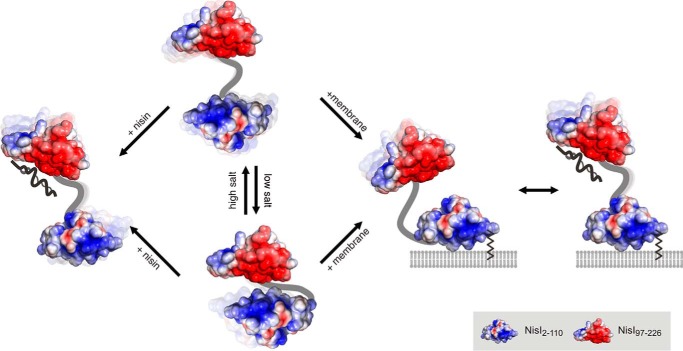
Many Gram-positive bacteria produce lantibiotics, genetically encoded and posttranslationally modified peptide antibiotics, which inhibit the growth of other Gram-positive bacteria. To protect themselves against their own lantibiotics these bacteria express a variety of immunity proteins including the LanI lipoproteins. The structural and mechanistic basis for LanI-mediated lantibiotic immunity is not yet understood. Lactococcus lactis produces the lantibiotic nisin, which is widely used as a food preservative. Its LanI protein NisI provides immunity against nisin but not against structurally very similar lantibiotics from other species such as subtilin from Bacillus subtilis. To understand the structural basis for LanI-mediated immunity and their specificity we investigated the structure of NisI. We found that NisI is a two-domain protein. Surprisingly, each of the two NisI domains has the same structure as the LanI protein from B. subtilis, SpaI, despite the lack of significant sequence homology. The two NisI domains and SpaI differ strongly in their surface properties and function. Additionally, SpaI-mediated lantibiotic immunity depends on the presence of a basic unstructured N-terminal region that tethers SpaI to the membrane. Such a region is absent from NisI. Instead, the N-terminal domain of NisI interacts with membranes but not with nisin. In contrast, the C-terminal domain specifically binds nisin and modulates the membrane affinity of the N-terminal domain. Thus, our results reveal an unexpected structural relationship between NisI and SpaI and shed light on the structural basis for LanI mediated lantibiotic immunity.
Keywords: antibiotic resistance; antibiotics; lantibiotic; lipoprotein; nisin binding; nuclear magnetic resonance (NMR); protein structure; small angle x-ray scattering.
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Klaenhammer T. R. (1988) Bacteriocins of lactic acid bacteria. Biochimie 70, 337–349 - PubMed
-
- Cotter P. D., Ross R. P., and Hill C. (2013) Bacteriocins: a viable alternative to antibiotics? Nat. Rev. Microbiol. 11, 95–105 - PubMed
-
- Dischinger J., Basi Chipalu S., and Bierbaum G. (2014) Lantibiotics: promising candidates for future applications in health care. Int. J. Med. Microbiol. 304, 51–62 - PubMed
-
- Arnison P. G., Bibb M. J., Bierbaum G., Bowers A. A., Bugni T. S., Bulaj G., Camarero J. A., Campopiano D. J., Challis G. L., Clardy J., Cotter P. D., Craik D. J., Dawson M., Dittmann E., Donadio S., Dorrestein P. C., Entian K.-D., Fischbach M. A., Garavelli J. S., Göransson U., Gruber C. W., Haft D. H., Hemscheidt T. K., Hertweck C., Hill C., Horswill A. R., Jaspars M., Kelly W. L., Klinman J. P., Kuipers O. P., Link A. J., Liu W., Marahiel M. A., Mitchell D. A., Moll G. N., Moore B. S., Müller R., Nair S. K., Nes I. F., Norris G. E., Olivera B. M., Onaka H., Patchett M. L., Piel J., Reaney M. J., Rebuffat S., Ross R. P., Sahl H.-G., Schmidt E. W., Selsted M. E., Severinov K., Shen B., Sivonen K., Smith L., Stein T., Süssmuth R. D., Tagg J. R., Tang G.-L., Truman A. W., Vederas J. C., Walsh C. T., Walton J. D., Wenzel S. C., Willey J. M., and van der Donk W. A. (2013) Ribosomally synthesized and post-translationally modified peptide natural products: overview and recommendations for a universal nomenclature. Nat. Prod. Rep. 30, 108–160 - PMC - PubMed
-
- Schnell N., Entian K. D., Schneider U., Götz F., Zähner H., Kellner R., and Jung G. (1988) Prepeptide sequence of epidermin, a ribosomally synthesized antibiotic with four sulphide-rings. Nature 333, 276–278 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
