

Caveolae protect endothelial cells from membrane rupture during increased cardiac output
- PMID: 26459598
- PMCID: PMC4602045
- DOI: 10.1083/jcb.201504042
Caveolae protect endothelial cells from membrane rupture during increased cardiac output
Abstract
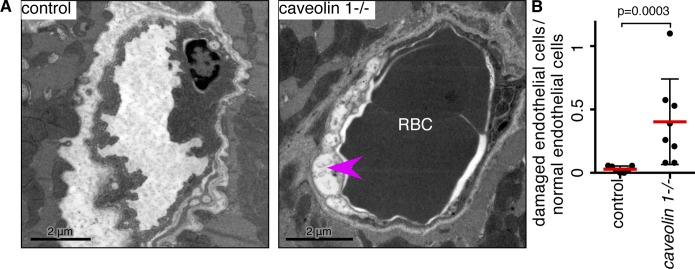
Caveolae are strikingly abundant in endothelial cells, yet the physiological functions of caveolae in endothelium and other tissues remain incompletely understood. Previous studies suggest a mechanoprotective role, but whether this is relevant under the mechanical forces experienced by endothelial cells in vivo is unclear. In this study we have sought to determine whether endothelial caveolae disassemble under increased hemodynamic forces, and whether caveolae help prevent acute rupture of the plasma membrane under these conditions. Experiments in cultured cells established biochemical assays for disassembly of caveolar protein complexes, and assays for acute loss of plasma membrane integrity. In vivo, we demonstrate that caveolae in endothelial cells of the lung and cardiac muscle disassemble in response to acute increases in cardiac output. Electron microscopy and two-photon imaging reveal that the plasma membrane of microvascular endothelial cells in caveolin 1(-/-) mice is much more susceptible to acute rupture when cardiac output is increased. These data imply that mechanoprotection through disassembly of caveolae is important for endothelial function in vivo.
© 2015 Cheng et al.
Figures
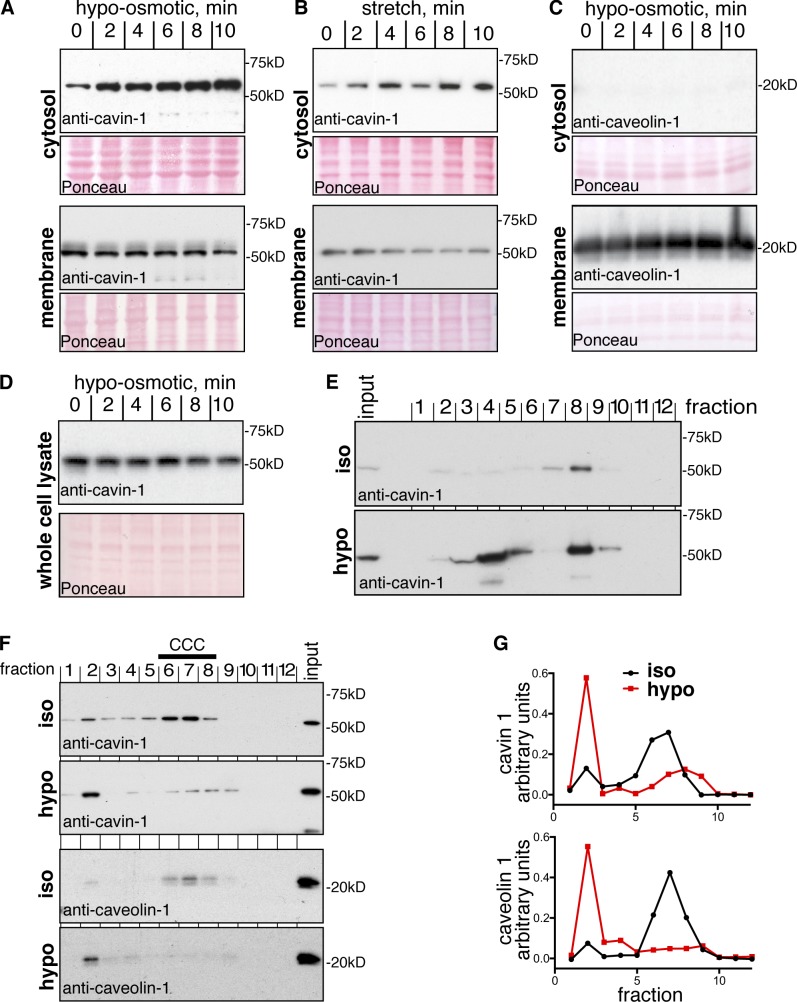
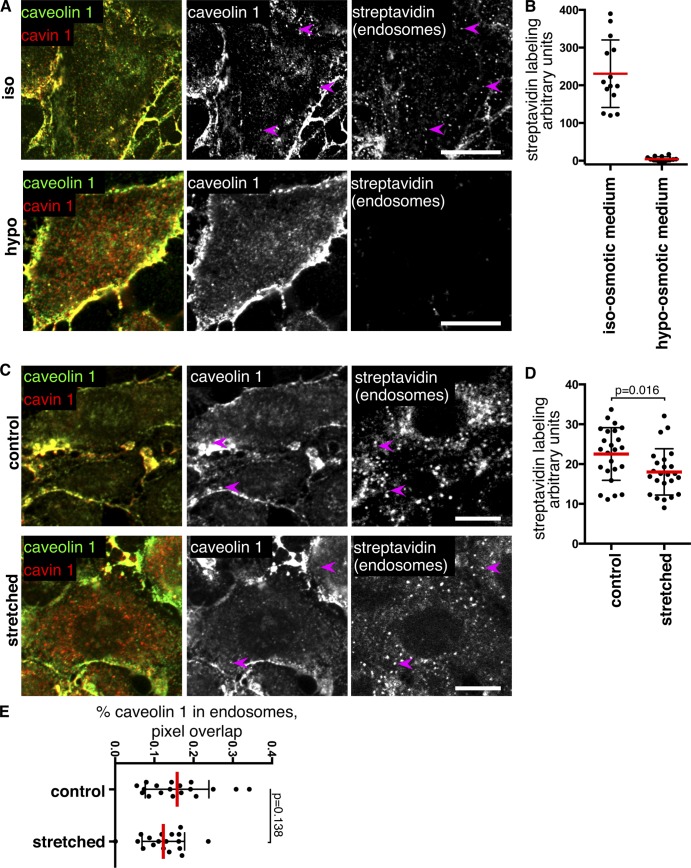
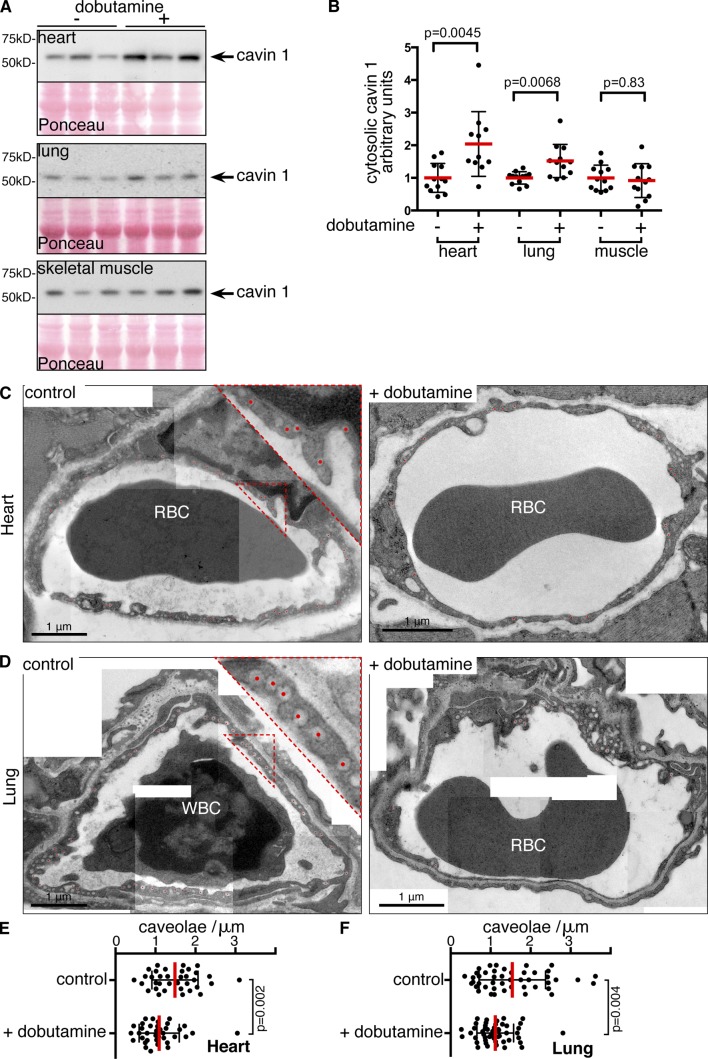
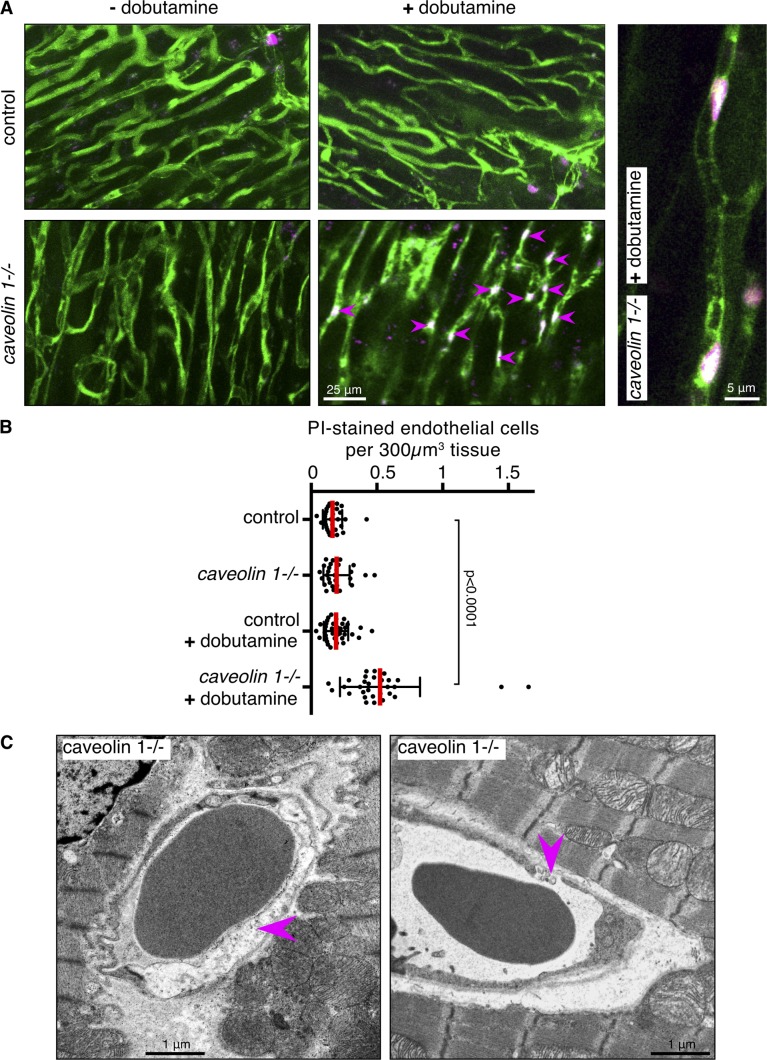
References
-
- Austin E.D., Ma L., LeDuc C., Berman Rosenzweig E., Borczuk A., Phillips J.A. III, Palomero T., Sumazin P., Kim H.R., Talati M.H., et al. . 2012. Whole exome sequencing to identify a novel gene (caveolin-1) associated with human pulmonary arterial hypertension. Circ Cardiovasc Genet. 5:336–343. 10.1161/CIRCGENETICS.111.961888 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
