

Evolutionary rescue by compensatory mutations is constrained by genomic and environmental backgrounds
- PMID: 26459777
- PMCID: PMC4631203
- DOI: 10.15252/msb.20156444
Evolutionary rescue by compensatory mutations is constrained by genomic and environmental backgrounds
Abstract
Since deleterious mutations may be rescued by secondary mutations during evolution, compensatory evolution could identify genetic solutions leading to therapeutic targets. Here, we tested this hypothesis and examined whether these solutions would be universal or would need to be adapted to one's genetic and environmental make-ups. We performed experimental evolutionary rescue in a yeast disease model for the Wiskott-Aldrich syndrome in two genetic backgrounds and carbon sources. We found that multiple aspects of the evolutionary rescue outcome depend on the genotype, the environment, or a combination thereof. Specifically, the compensatory mutation rate and type, the molecular rescue mechanism, the genetic target, and the associated fitness cost varied across contexts. The course of compensatory evolution is therefore highly contingent on the initial conditions in which the deleterious mutation occurs. In addition, these results reveal biologically favored therapeutic targets for the Wiskott-Aldrich syndrome, including the target of an unrelated clinically approved drug. Our results experimentally illustrate the importance of epistasis and environmental evolutionary constraints that shape the adaptive landscape and evolutionary rate of molecular networks.
Keywords: Wiskott–Aldrich syndrome; aneuploidy; epistasis; experimental evolution; genotype‐by‐environment interaction.
© 2015 The Authors. Published under the terms of the CC BY 4.0 license.
Figures



Compensatory mutation rate of the thermosensitive las17-41 phenotype in four contexts: RM and BY genetic backgrounds and two carbon sources, glucose (Glu) and galactose (Gal) (left axis, n = 3, 8, 4, and 4 for BY gal, BY glu, RM gal, and RM glu, respectively). Bars = average, error bars = SD. Overlaid dots show the number of unique rescue mutation (right axis) found in each context. Mutations at a shared locus and nucleotide position were considered equivalent.
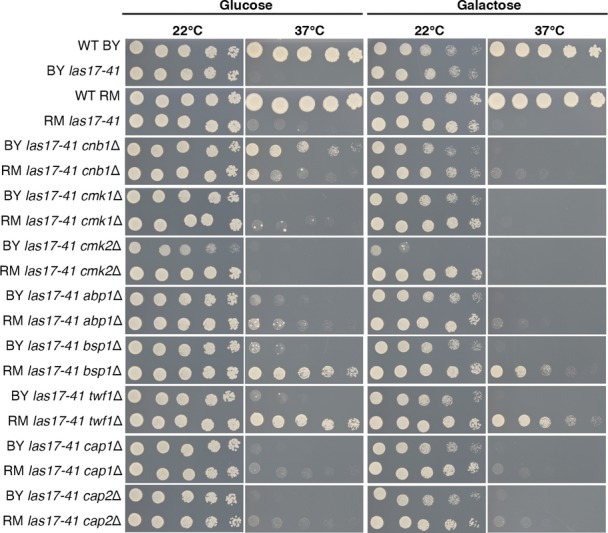
The average growth of evolved strains sharing compensatory mutation types at 22°C (top panel) and 37°C (bottom panel) in four culture media is shown relative to their respective progenitor. The las17 strains carry R41W, R41L, or R41Q functional reversions. WT = wild-type strain. Growth phenotypes were measured on synthetic complete (SC) and rich media (YP), n = number of strains next to labels, each inferred from eight replicates. One cap1 mutant was excluded because of mitochondrial loss and one mutant because it contained two rescue mutations (act1 and cap2).
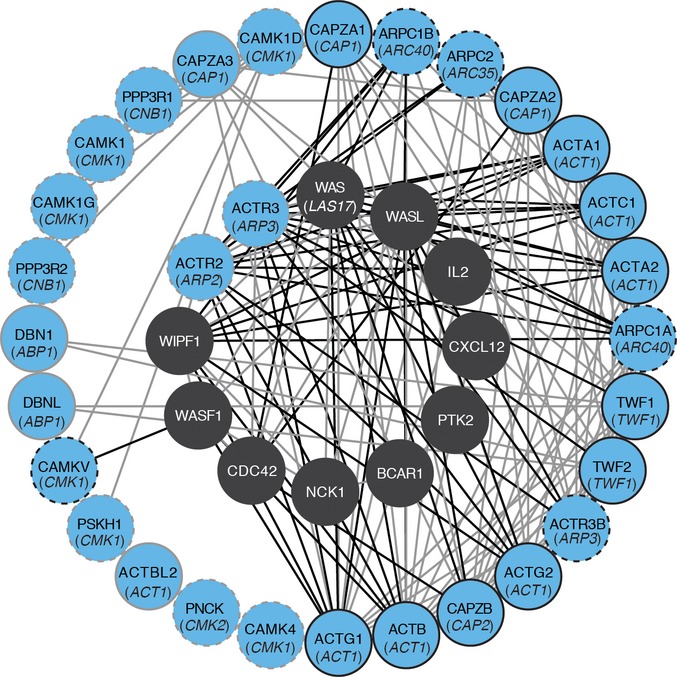
Proportion of compensatory mutations identified in the four experimental contexts. Only two genes were identified in all four contexts, indicating that at the target level, up to 80% of the mutations could be context dependent.
Proportion of each rescue mutation type encountered in each experiment, n = same as in C, SNV = single nucleotide variant.
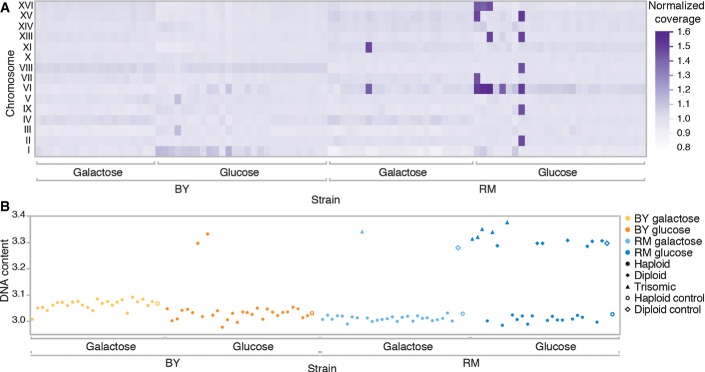
Six strains had unequal read coverage across chromosomes, with a coverage of approximately 1.5× along up to six entire chromosomes. This factor is consistent with trisomia in diploid strains. The heat map shows the normalized average coverage across chromosomes for each sequenced mutant strains. ChXII is not shown because of a constant bias in its coverage estimate caused by the multiple copies of rRNA genes it encodes.
Ploidy measurements of all the sequenced strains using DNA content measurement by flow cytometry confirmed trisomia. Haploid and diploid assignments were determined by k-means clustering on test and control strains. Trisomic assignment is based on the unequal coverage values in (A). A diploid control strain of the RM genetic background is also included. Some strains for which a coding mutation was identified were also diploid. Because the frequency of the variant was in all cases near 100% (Table EV1), meaning that the diploid mutants are homozygous for the rescue mutation, it can be assumed that diploidization occurred after the mutation.


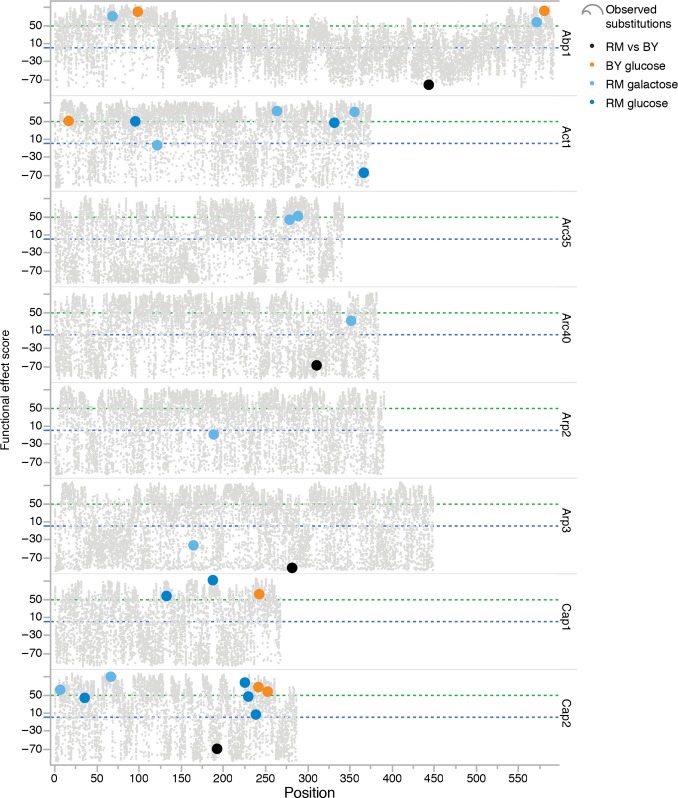
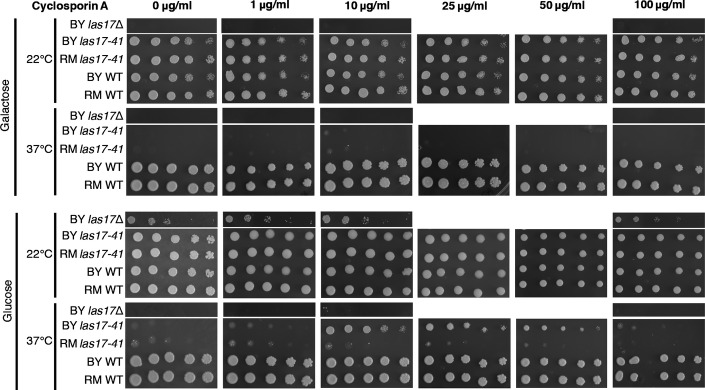
Bars show the fraction of colonies transformed with a mutant allele showing more growth at 37°C than 95% of the control, that is, the originating wild-type allele in BY and RM las17-41 thermosensitive strains on glucose and galactose. The x-axis indicates the amino acid changes for each allele. A low fraction of compensated transformants could reflect that the addition of the mutant allele increases the compensatory mutation rate only or that it has a partially recessive effect. Because the wild-type copy is also present in the transformed haploid strains, a recessive gain of function would not be revealed by this experiment.
Increasing the gene dosage of actin by adding a wild-type actin allele from BY or RM (x-axis) can compensate las17-41 thermosensitivity in the RM background on glucose. Bars show the fraction of colonies that grow more than 95% of control colonies at 37°C. Control colonies are BY and RM las17-41 transformed with an empty plasmid (pRS316), which reflects the background growth owed to compensatory mutants arising during the experiment.

References
-
- Albert MH, Notarangelo LD, Ochs HD. Clinical spectrum, pathophysiology and treatment of the Wiskott–Aldrich syndrome. Curr Opin Hematol. 2011;18:42–48. - PubMed
-
- Ashworth A, Lord CJ, Reis-Filho JS. Genetic interactions in cancer progression and treatment. Cell. 2011;145:30–38. - PubMed
-
- Bandyopadhyay S, Mehta M, Kuo D, Sung MK, Chuang R, Jaehnig EJ, Bodenmiller B, Licon K, Copeland W, Shales M, Fiedler D, Dutkowski J, Guénolé A, van Attikum H, Shokat KM, Kolodner RD, Huh WK, Aebersold R, Keogh MC, Krogan NJ, et al. Rewiring of genetic networks in response to DNA damage. Science. 2010;330:1385–1389. . - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
