

Bursts of beta oscillation differentiate postperformance activity in the striatum and motor cortex of monkeys performing movement tasks
- PMID: 26460033
- PMCID: PMC4640760
- DOI: 10.1073/pnas.1517629112
Bursts of beta oscillation differentiate postperformance activity in the striatum and motor cortex of monkeys performing movement tasks
Abstract
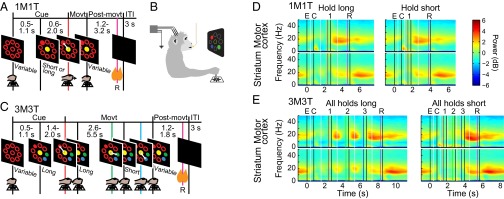
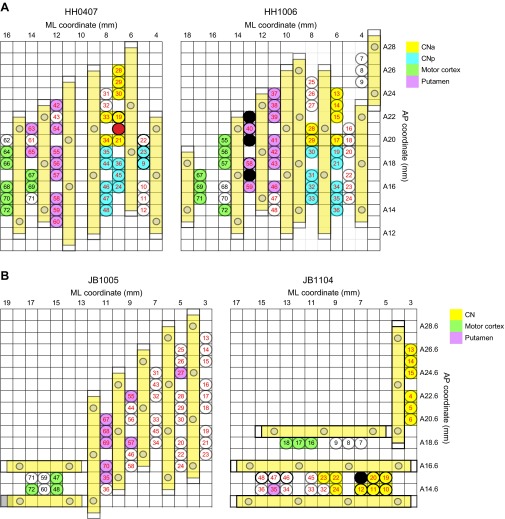
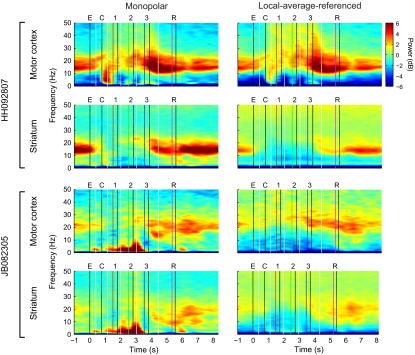
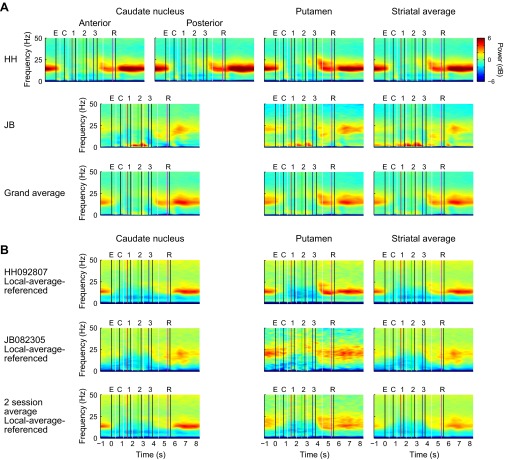
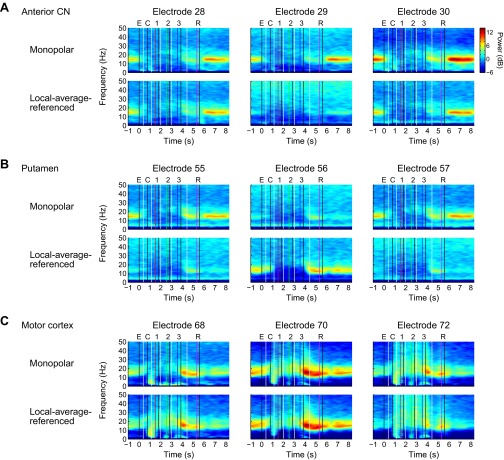
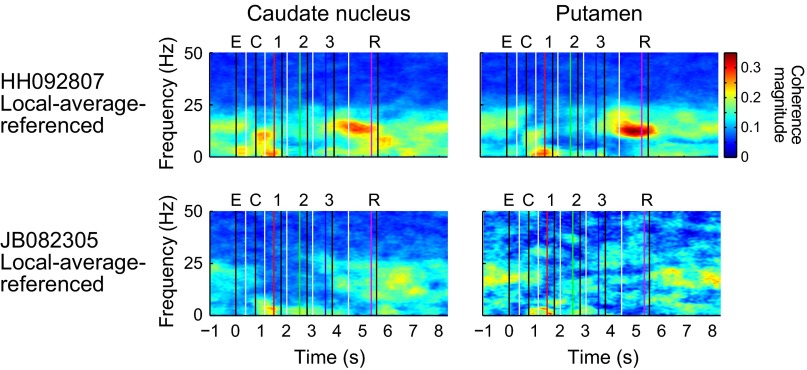
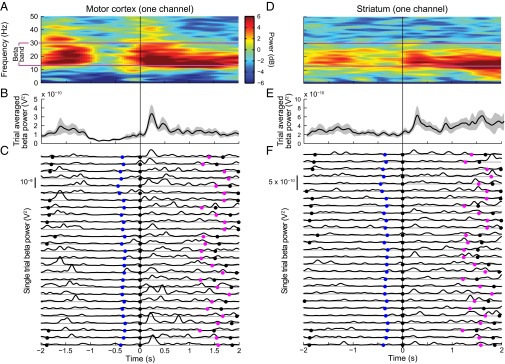
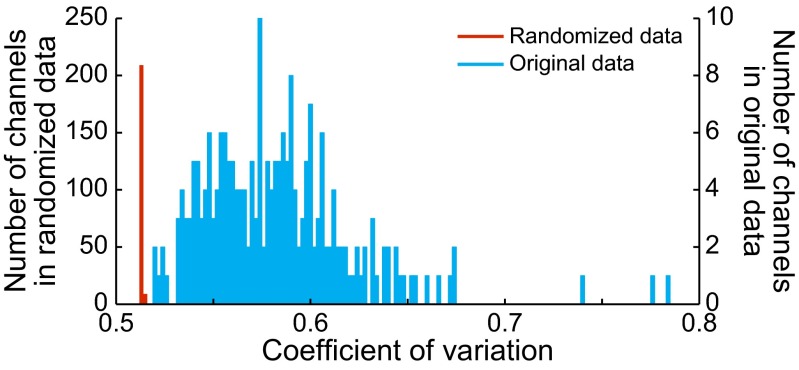
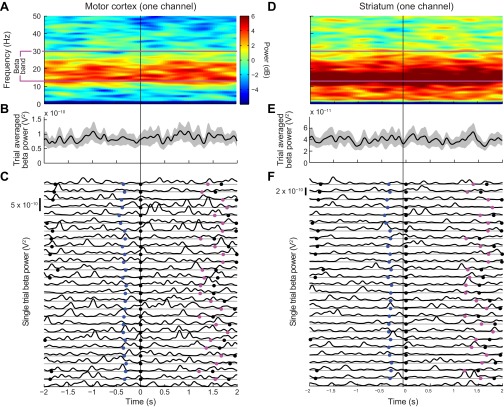
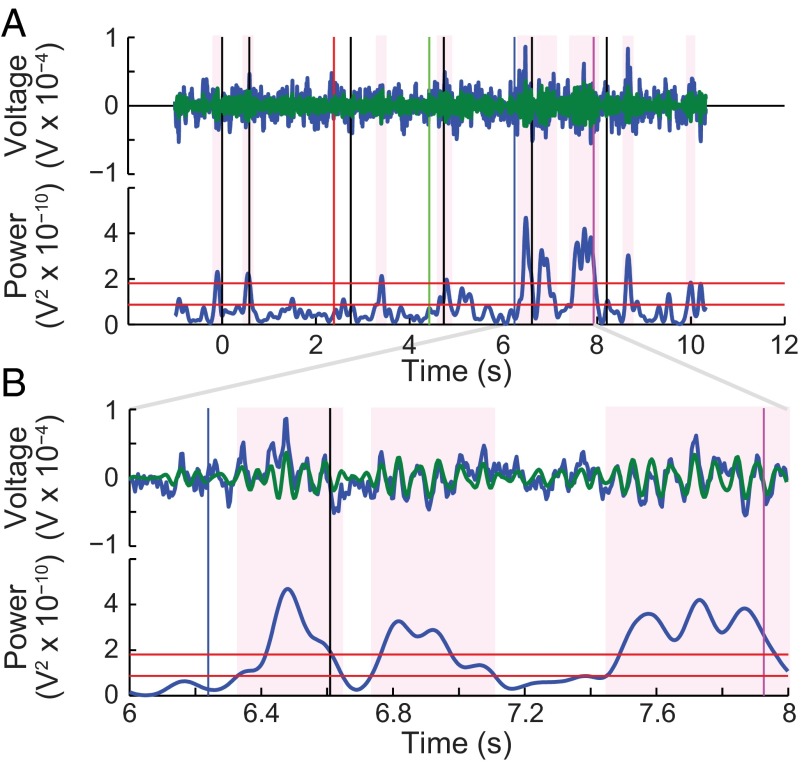
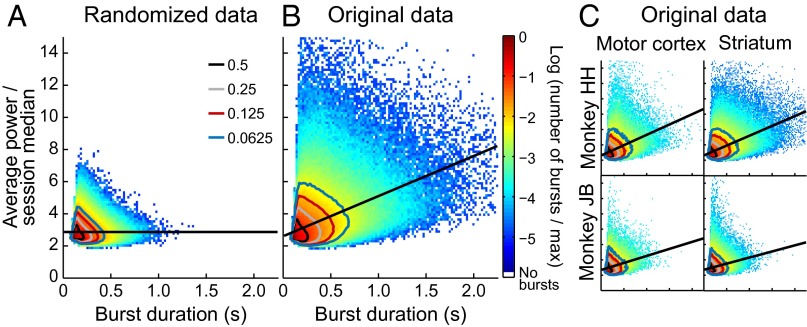
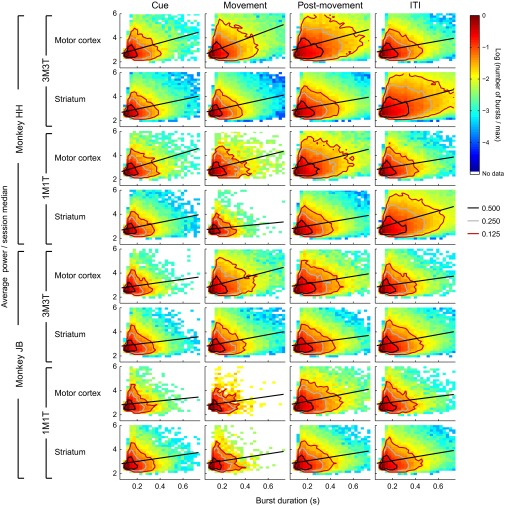
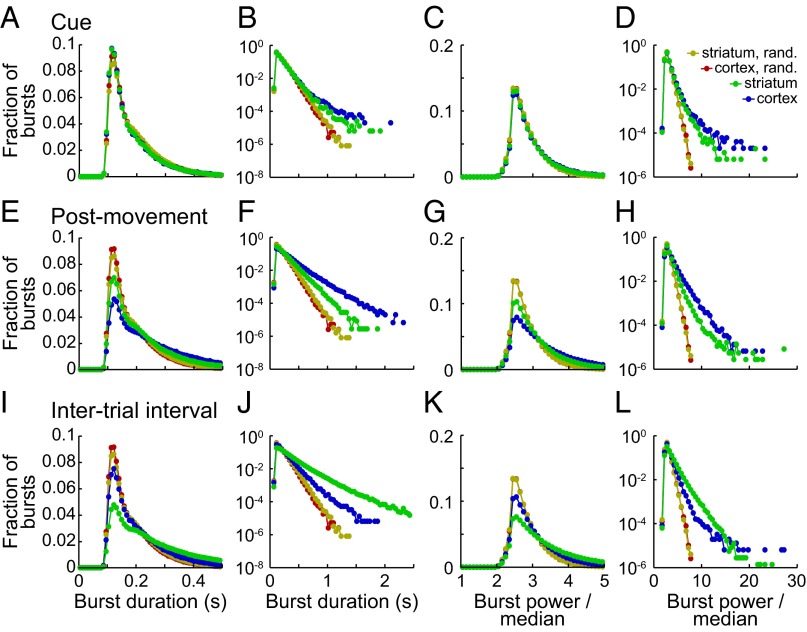
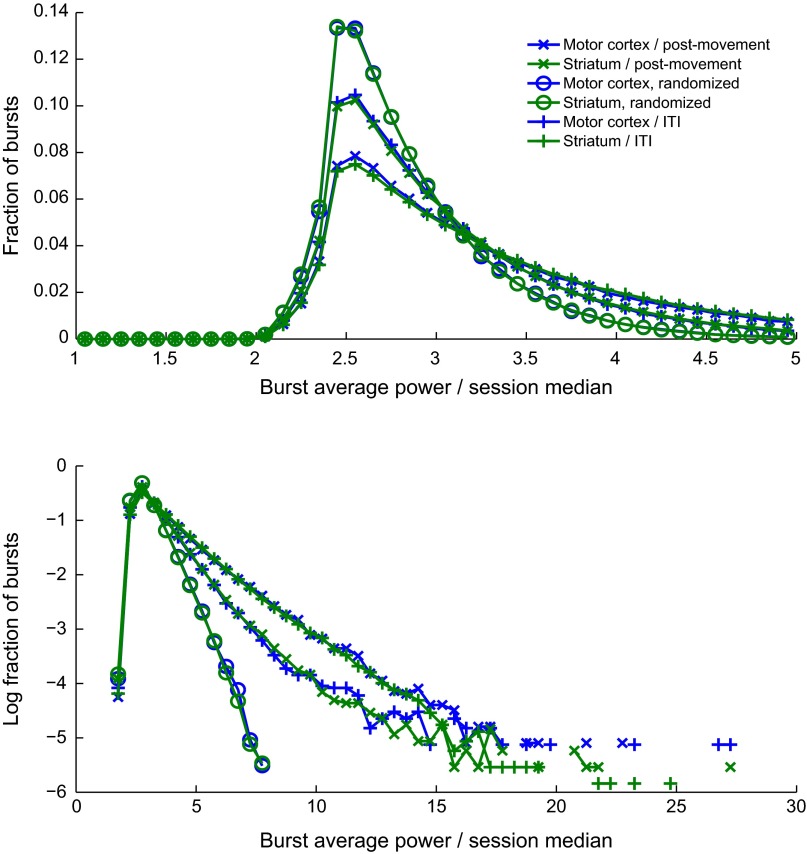
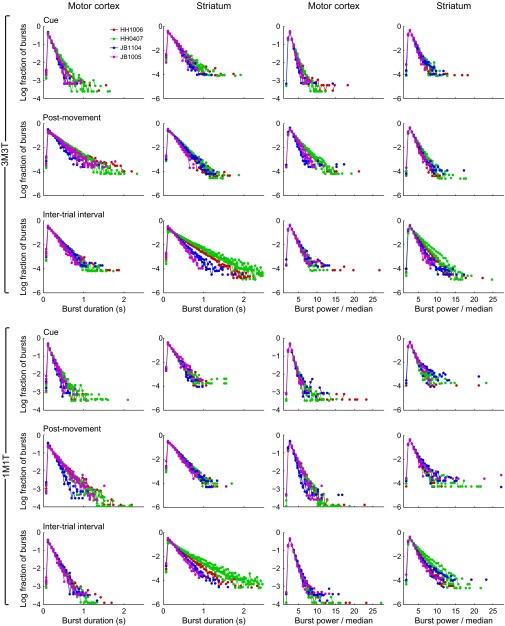
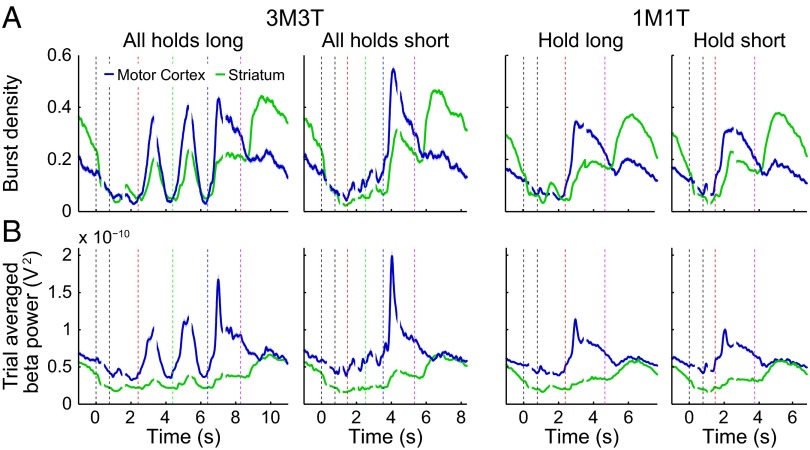
Studies of neural oscillations in the beta band (13-30 Hz) have demonstrated modulations in beta-band power associated with sensory and motor events on time scales of 1 s or more, and have shown that these are exaggerated in Parkinson's disease. However, even early reports of beta activity noted extremely fleeting episodes of beta-band oscillation lasting <150 ms. Because the interpretation of possible functions for beta-band oscillations depends strongly on the time scale over which they occur, and because of these oscillations' potential importance in Parkinson's disease and related disorders, we analyzed in detail the distributions of duration and power for beta-band activity in a large dataset recorded in the striatum and motor-premotor cortex of macaque monkeys performing reaching tasks. Both regions exhibited typical beta-band suppression during movement and postmovement rebounds of up to 3 s as viewed in data averaged across trials, but single-trial analysis showed that most beta oscillations occurred in brief bursts, commonly 90-115 ms long. In the motor cortex, the burst probabilities peaked following the last movement, but in the striatum, the burst probabilities peaked at task end, after reward, and continued through the postperformance period. Thus, what appear to be extended periods of postperformance beta-band synchronization reflect primarily the modulated densities of short bursts of synchrony occurring in region-specific and task-time-specific patterns. We suggest that these short-time-scale events likely underlie the functions of most beta-band activity, so that prolongation of these beta episodes, as observed in Parkinson's disease, could produce deleterious network-level signaling.
Keywords: basal ganglia; beta band; local field potentials; sequential movement; synchronization.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Neuper C, Wortz M, Pfurtscheller G. ERD/ERS patterns reflecting sensorimotor activation and deactivation. Prog Brain Res. 2006;159:211–222. - PubMed
-
- Stancak A, Jr, Pfurtscheller G. Event-related desynchronisation of central beta-rhythms during brisk and slow self-paced finger movements of dominant and nondominant hand. Brain Res Cogn Brain Res. 1996;4(3):171–183. - PubMed
-
- Rubino D, Robbins KA, Hatsopoulos NG. Propagating waves mediate information transfer in the motor cortex. Nat Neurosci. 2006;9(12):1549–1557. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
