

Adult Lineage-Restricted CNS Progenitors Specify Distinct Glioblastoma Subtypes
- PMID: 26461091
- PMCID: PMC4607935
- DOI: 10.1016/j.ccell.2015.09.007
Adult Lineage-Restricted CNS Progenitors Specify Distinct Glioblastoma Subtypes
Abstract
A central question in glioblastoma multiforme (GBM) research is the identity of the tumor-initiating cell, and its contribution to the malignant phenotype and genomic state. We examine the potential of adult lineage-restricted progenitors to induce fully penetrant GBM using CNS progenitor-specific inducible Cre mice to mutate Nf1, Trp53, and Pten. We identify two phenotypically and molecularly distinct GBM subtypes governed by identical driver mutations. We demonstrate that the two subtypes arise from functionally independent pools of adult CNS progenitors. Despite histologic identity as GBM, these tumor types are separable based on the lineage of the tumor-initiating cell. These studies point to the cell of origin as a major determinant of GBM subtype diversity.
Copyright © 2015 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors report no conflict of interest.
Figures
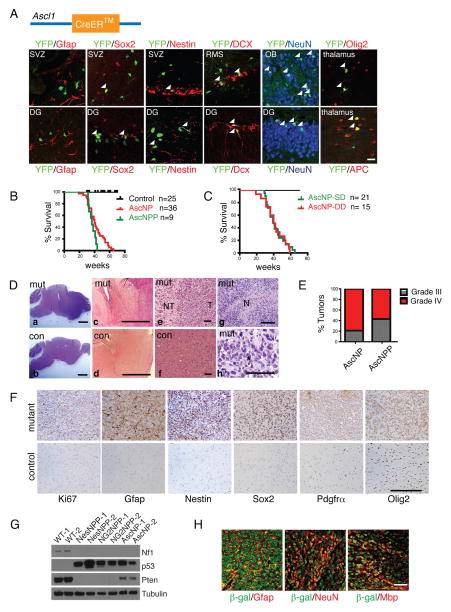
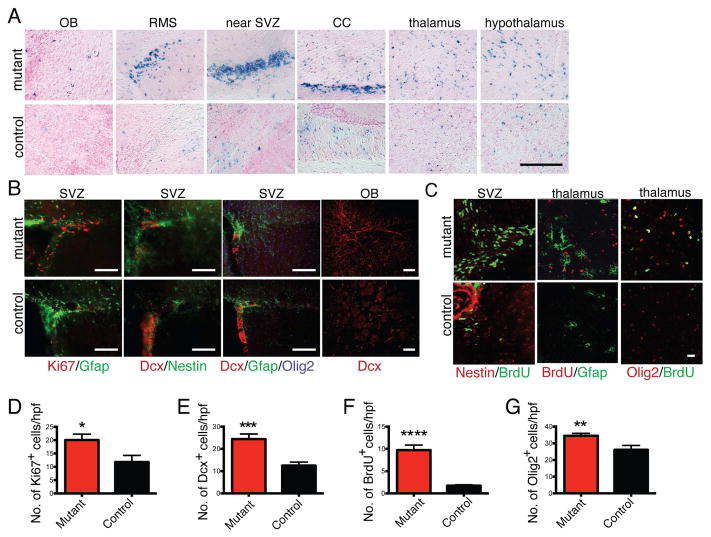
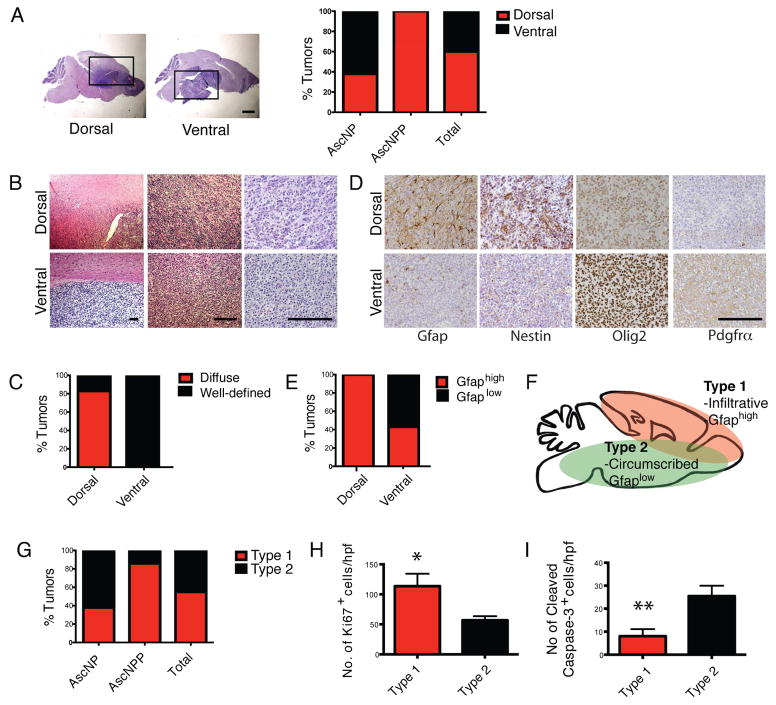
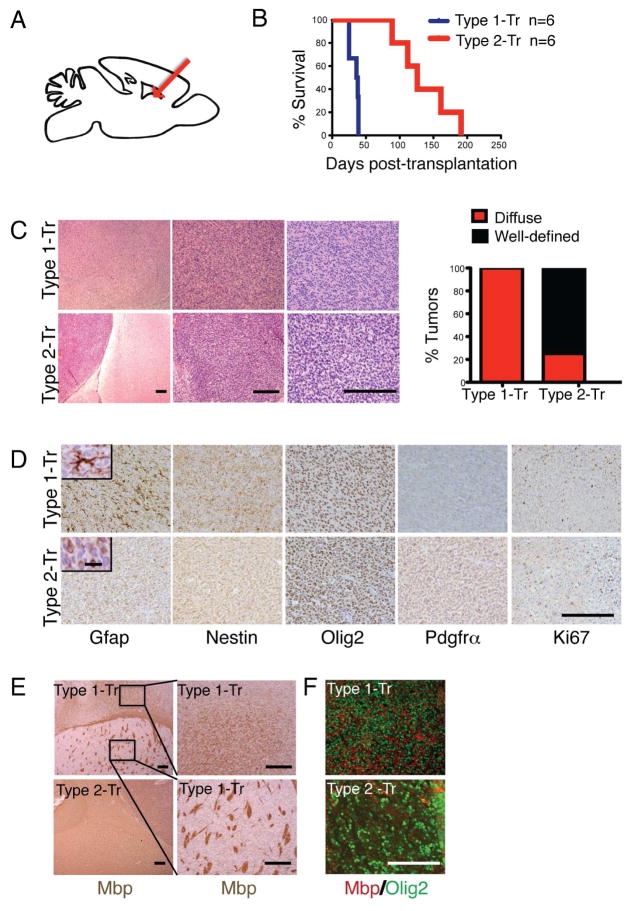
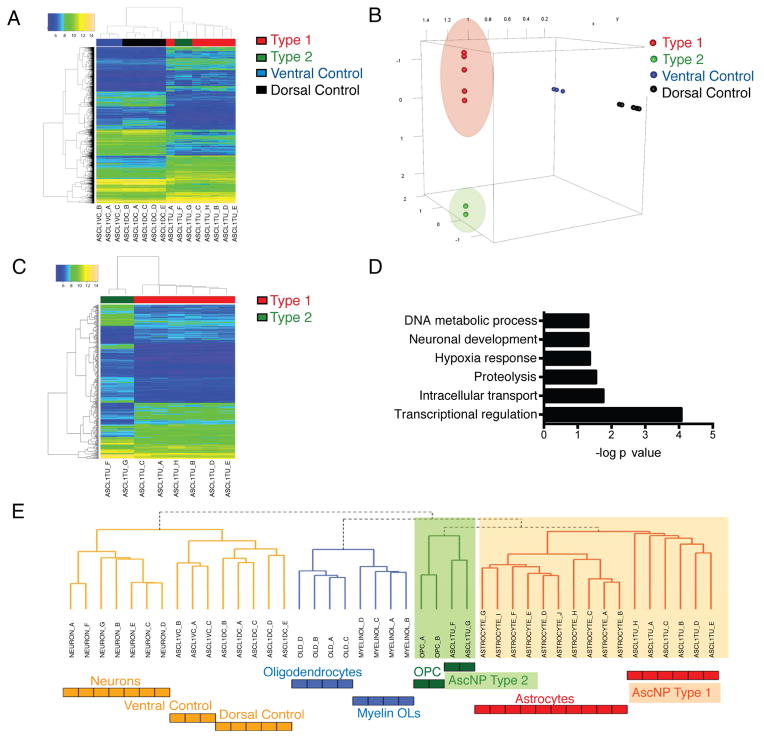
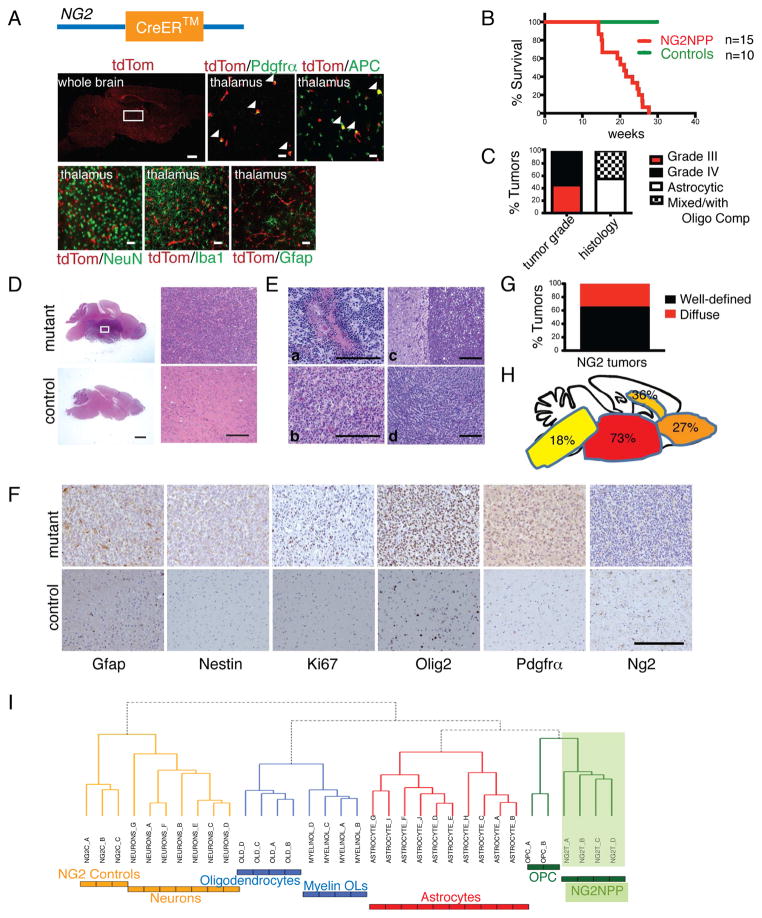
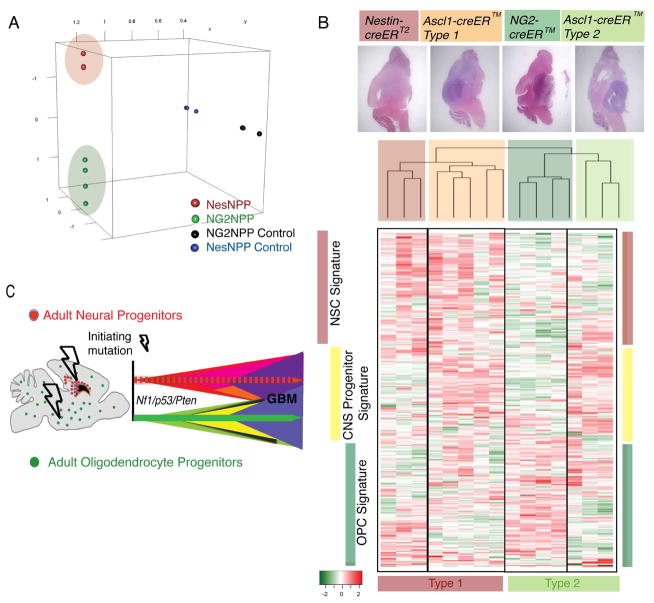
Comment in
-
Of Brains and Blood: Developmental Origins of Glioma Diversity?Cancer Cell. 2015 Oct 12;28(4):403-404. doi: 10.1016/j.ccell.2015.09.013. Cancer Cell. 2015. PMID: 26461085
References
-
- Alvarez-Buylla A, Lim DA. For the long run: maintaining germinal niches in the adult brain. Neuron. 2004;41:683–686. - PubMed
-
- Beckervordersandforth R, Tripathi P, Ninkovic J, Bayam E, Lepier A, Stempfhuber B, Kirchhoff F, Hirrlinger J, Haslinger A, Lie DC, et al. In vivo fate mapping and expression analysis reveals molecular hallmarks of prospectively isolated adult neural stem cells. Cell Stem Cell. 2010;7:744–758. - PubMed
-
- Cahoy JD, Emery B, Kaushal A, Foo LC, Zamanian JL, Christopherson KS, Xing Y, Lubischer JL, Krieg PA, Krupenko SA, et al. A transcriptome database for astrocytes, neurons, and oligodendrocytes: a new resource for understanding brain development and function. J Neurosci. 2008;28:264–278. - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
