

Transgenic rescue of phenotypic deficits in a mouse model of alternating hemiplegia of childhood
- PMID: 26463346
- PMCID: PMC4701769
- DOI: 10.1007/s10048-015-0461-1
Transgenic rescue of phenotypic deficits in a mouse model of alternating hemiplegia of childhood
Abstract
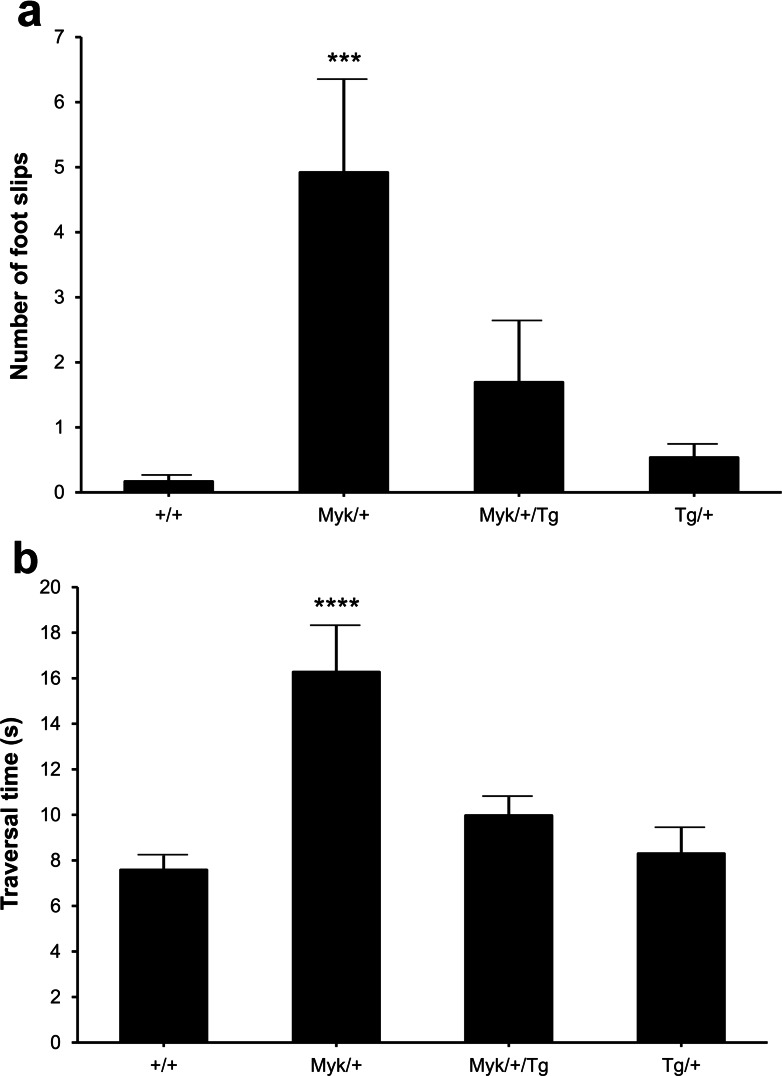
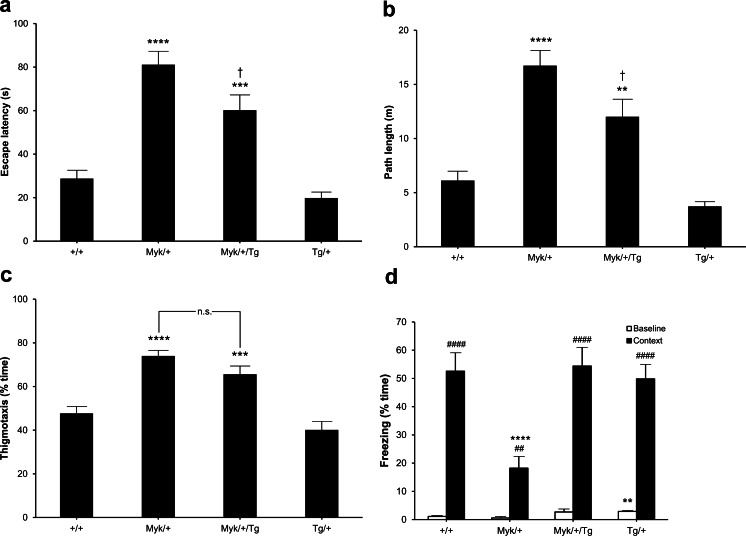
Missense mutations in ATP1A3 encoding Na(+),K(+)-ATPase α3 are the primary cause of alternating hemiplegia of childhood (AHC). Most ATP1A3 mutations in AHC lie within a cluster in or near transmembrane α-helix TM6, including I810N that is also found in the Myshkin mouse model of AHC. These mutations all substantially reduce Na(+),K(+)-ATPase α3 activity. Herein, we show that Myshkin mice carrying a wild-type Atp1a3 transgene that confers a 16 % increase in brain-specific total Na(+),K(+)-ATPase activity show significant phenotypic improvements compared with non-transgenic Myshkin mice. Interventions to increase the activity of wild-type Na(+),K(+)-ATPase α3 in AHC patients should be investigated further.
Keywords: Alternating hemiplegia; Atp1a3; Mice; Na+,K+-ATPase α3; Transgenic rescue.
Figures

References
-
- Sweney MT, Silver K, Gerard-Blanluet M, Pedespan JM, Renault F, Arzimanoglou A, Schlesinger-Massart M, Lewelt AJ, Reyna SP, Swoboda KJ. Alternating hemiplegia of childhood: early characteristics and evolution of a neurodevelopmental syndrome. Pediatrics. 2009;123:e534–41. doi: 10.1542/peds.2008-2027. - DOI - PubMed
-
- Panagiotakaki E, Gobbi G, Neville B, Ebinger F, Campistol J, Nevsímalová S, Laan L, Casaer P, Spiel G, Giannotta M, Fons C, Ninan M, Sange G, Schyns T, Vavassori R, Poncelin D, ENRAH Consortium. Arzimanoglou A. Evidence of a non-progressive course of alternating hemiplegia of childhood: study of a large cohort of children and adults. Brain. 2010;133:3598–610. doi: 10.1093/brain/awq295. - DOI - PubMed
-
- Heinzen EL, Arzimanoglou A, Brashear A, Clapcote SJ, Gurrieri F, Goldstein DB, Jóhannesson SH, Mikati MA, Neville B, Nicole S, Ozelius LJ, Poulsen H, Schyns T, Sweadner KJ, van den Maagdenberg A, Vilsen B, ATP1A3 Working Group Distinct neurological disorders with ATP1A3 mutations. Lancet Neurol. 2014;13:503–14. doi: 10.1016/S1474-4422(14)70011-0. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
