

Developmental Function of the PHR Protein RPM-1 Is Required for Learning in Caenorhabditis elegans
- PMID: 26464359
- PMCID: PMC4683646
- DOI: 10.1534/g3.115.021410
Developmental Function of the PHR Protein RPM-1 Is Required for Learning in Caenorhabditis elegans
Abstract
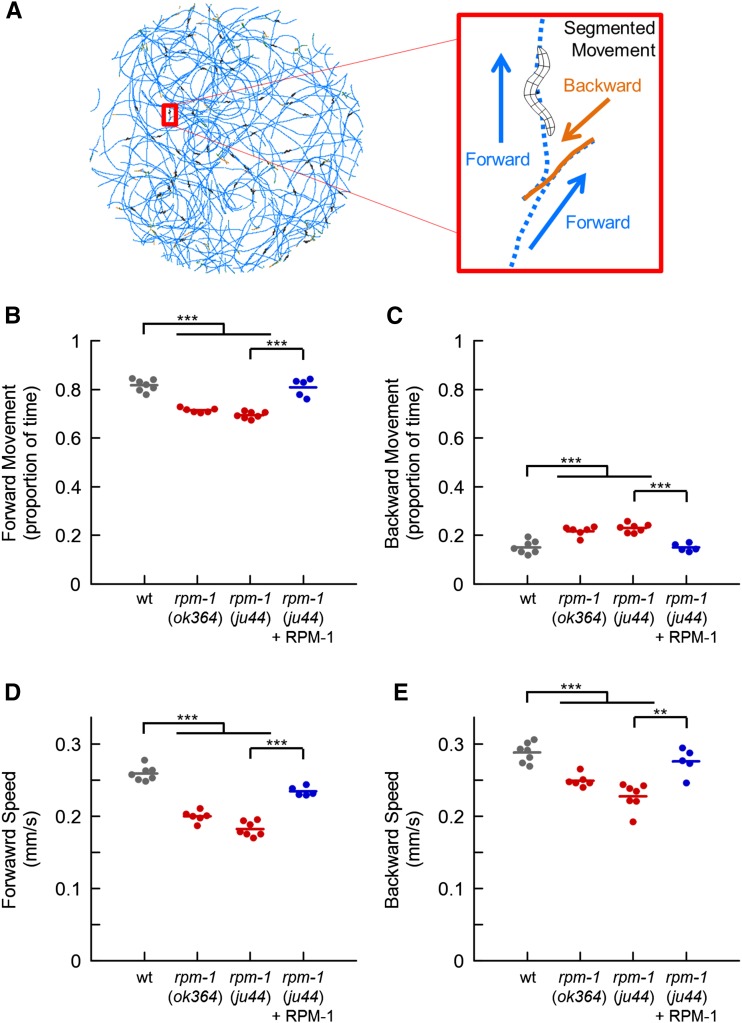
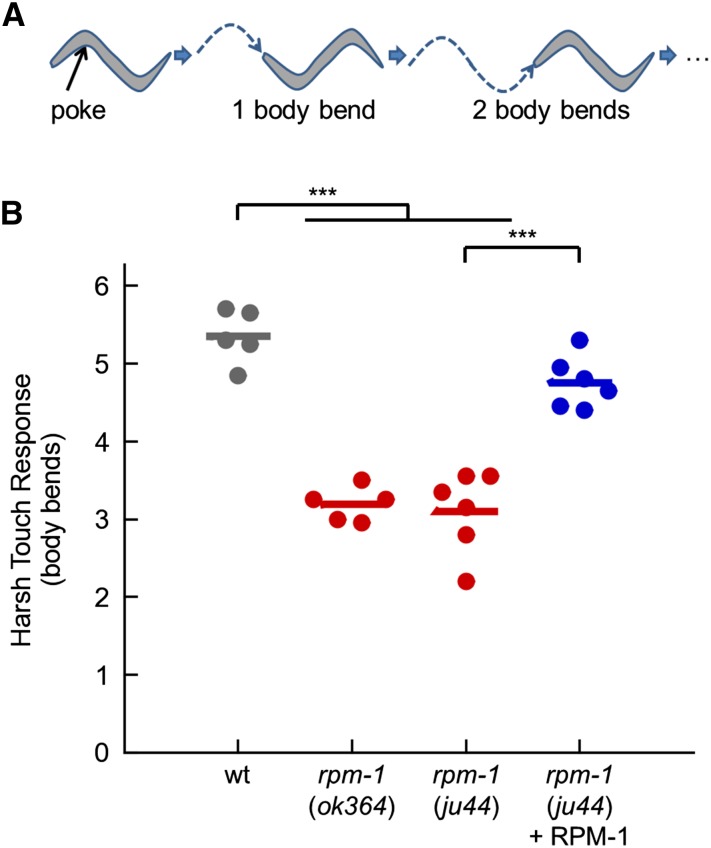
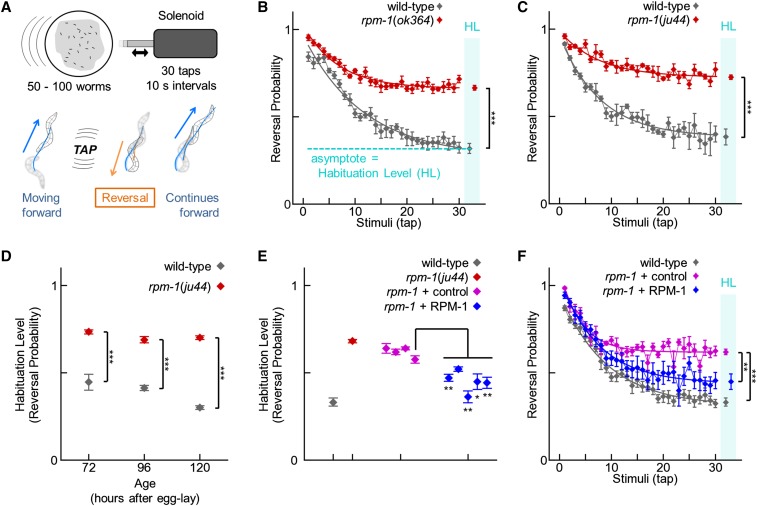
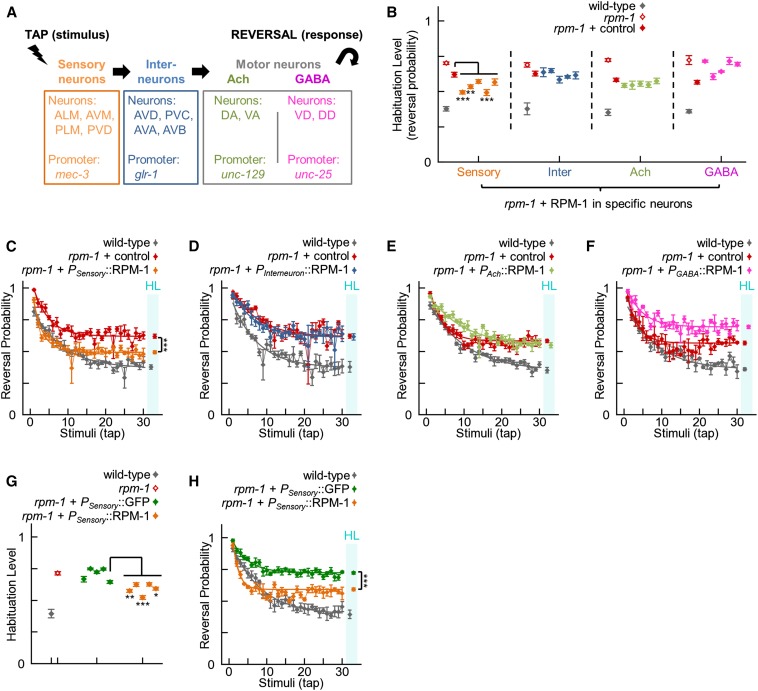
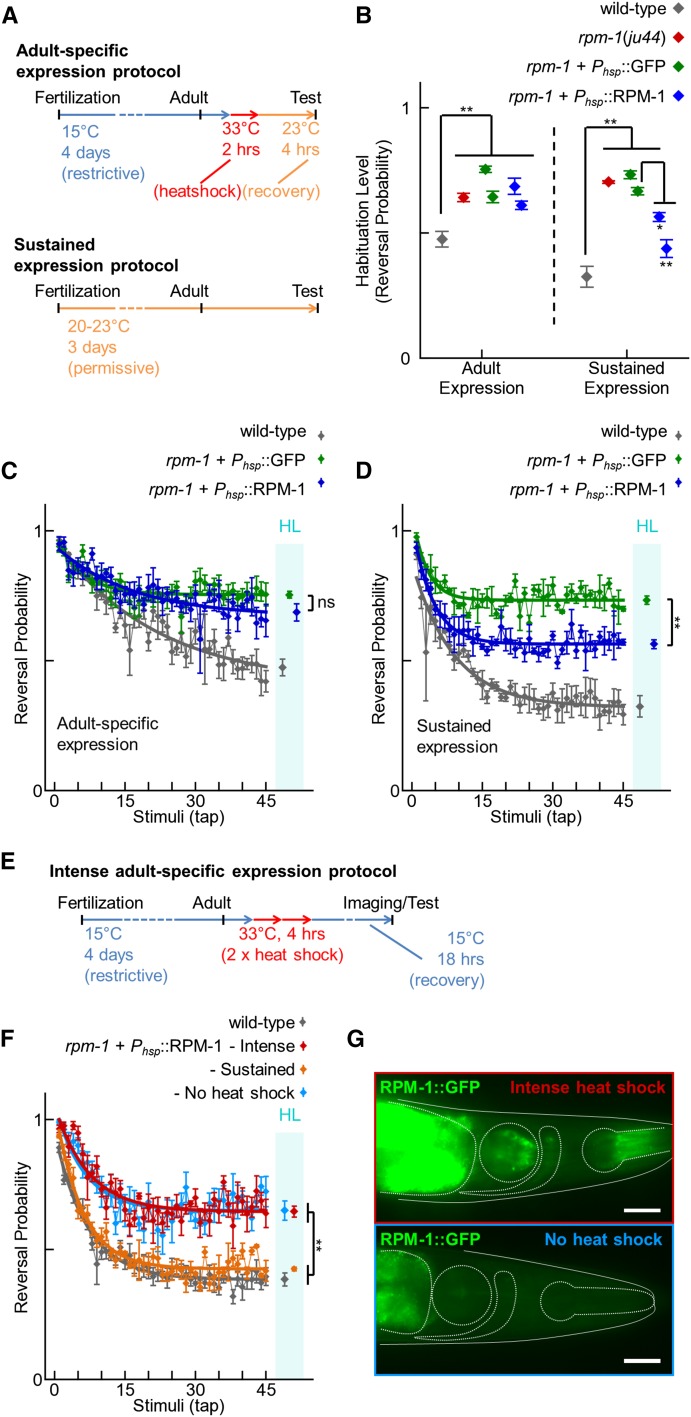
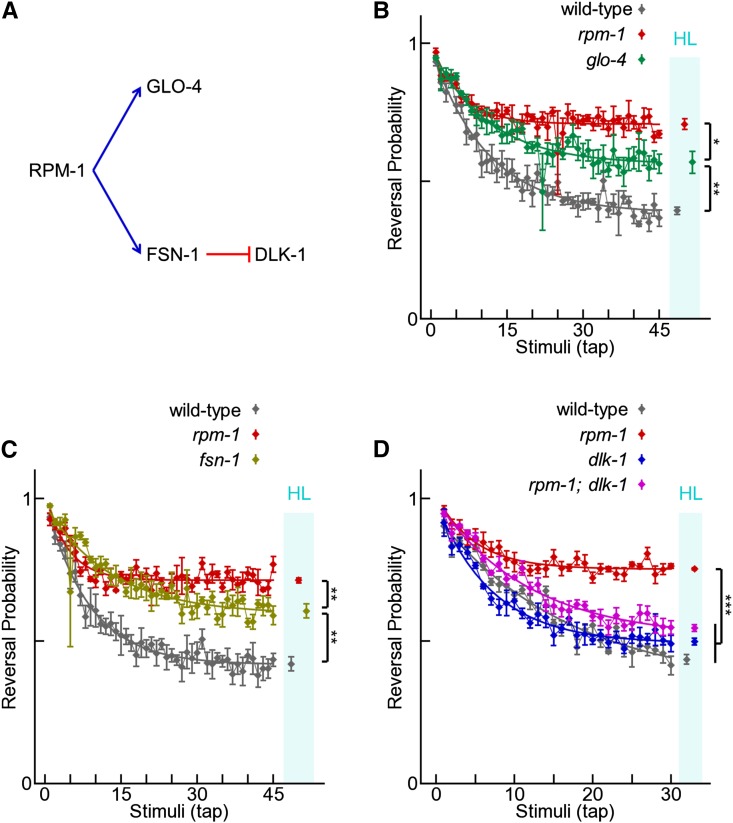
The PAM/Highwire/RPM-1 (PHR) proteins are signaling hubs that function as important regulators of neural development. Loss of function in Caenorhabditis elegans rpm-1 and Drosophila Highwire results in failed axon termination, inappropriate axon targeting, and abnormal synapse formation. Despite broad expression in the nervous system and relatively dramatic defects in synapse formation and axon development, very mild abnormalities in behavior have been found in animals lacking PHR protein function. Therefore, we hypothesized that large defects in behavior might only be detected in scenarios in which evoked, prolonged circuit function is required, or in which behavioral plasticity occurs. Using quantitative approaches in C. elegans, we found that rpm-1 loss-of-function mutants have relatively mild abnormalities in exploratory locomotion, but have large defects in evoked responses to harsh touch and learning associated with tap habituation. We explored the nature of the severe habituation defects in rpm-1 mutants further. To address what part of the habituation circuit was impaired in rpm-1 mutants, we performed rescue analysis with promoters for different neurons. Our findings indicate that RPM-1 function in the mechanosensory neurons affects habituation. Transgenic expression of RPM-1 in adult animals failed to rescue habituation defects, consistent with developmental defects in rpm-1 mutants resulting in impaired habituation. Genetic analysis showed that other regulators of neuronal development that function in the rpm-1 pathway (including glo-4, fsn-1, and dlk-1) also affected habituation. Overall, our findings suggest that developmental defects in rpm-1 mutants manifest most prominently in behaviors that require protracted or plastic circuit function, such as learning.
Keywords: C. elegans; PHR protein; RPM-1; habituation; learning.
Copyright © 2015 Giles et al.
Figures
References
-
- Ardiel E. L., Rankin C. H., 2010. An elegant mind: learning and memory in Caenorhabditis elegans. Learn. Mem. 17: 191–201. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous