

Explaining the striking difference in twist-stretch coupling between DNA and RNA: A comparative molecular dynamics analysis
- PMID: 26464435
- PMCID: PMC4666353
- DOI: 10.1093/nar/gkv1028
Explaining the striking difference in twist-stretch coupling between DNA and RNA: A comparative molecular dynamics analysis
Abstract
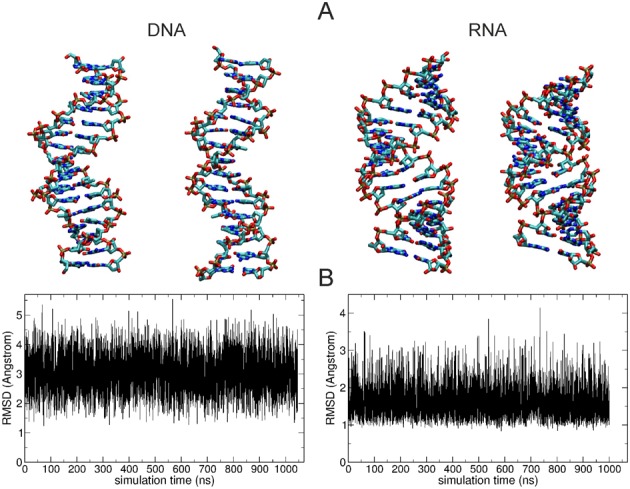
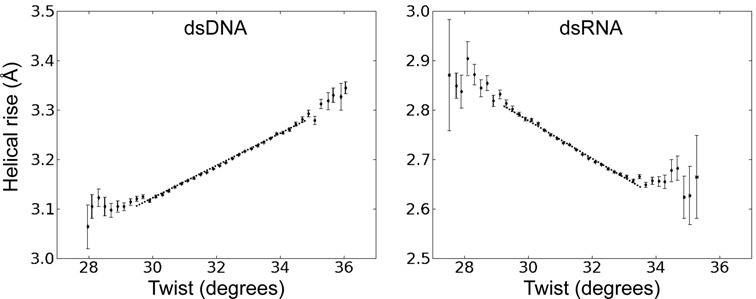
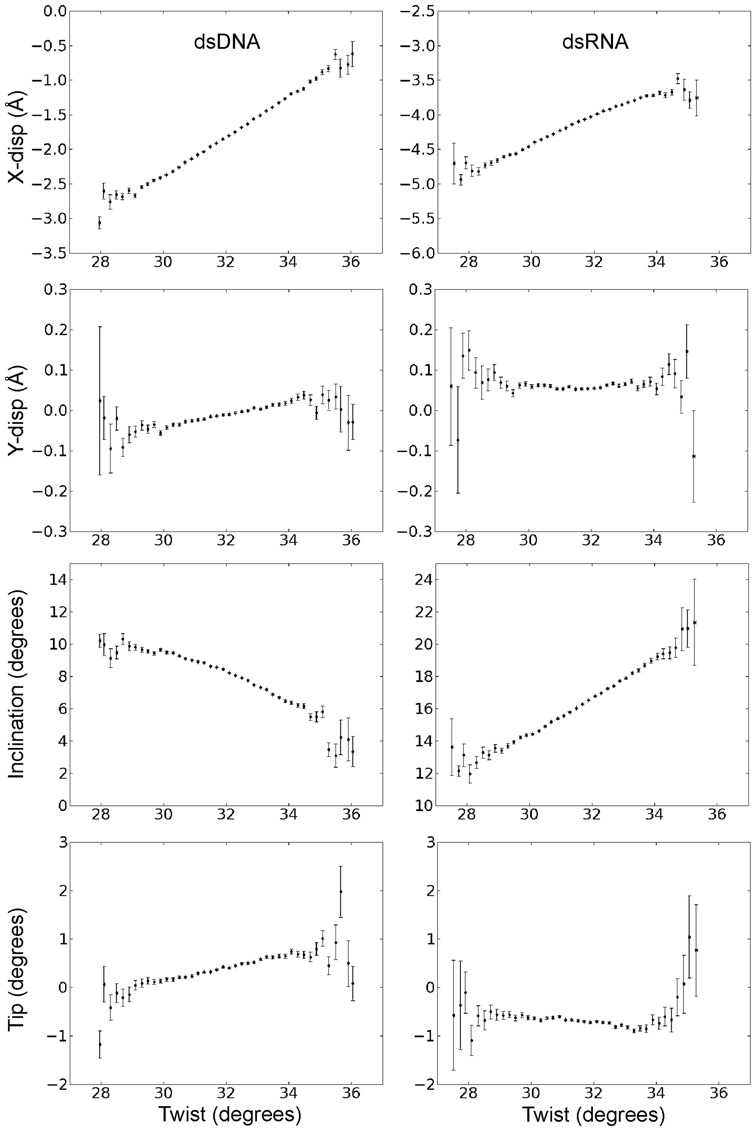
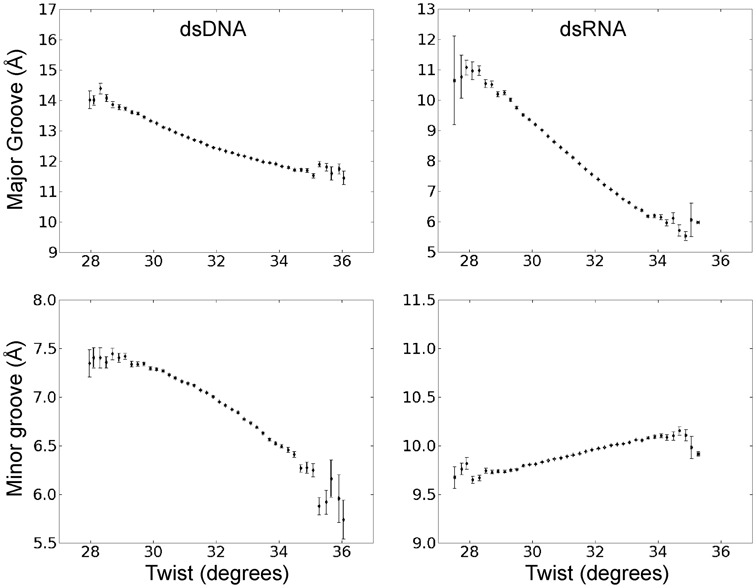
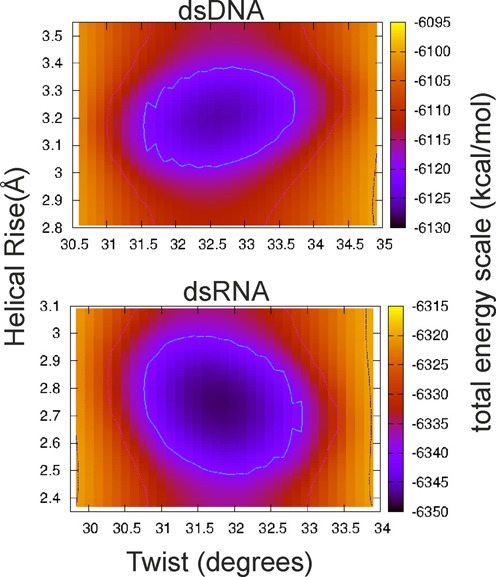
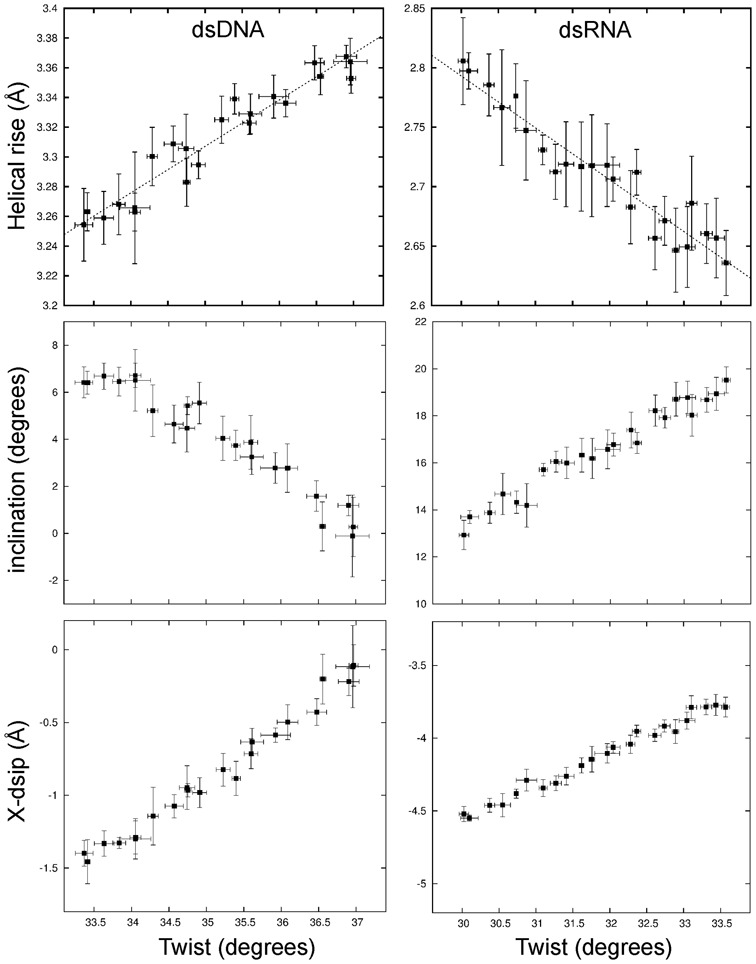
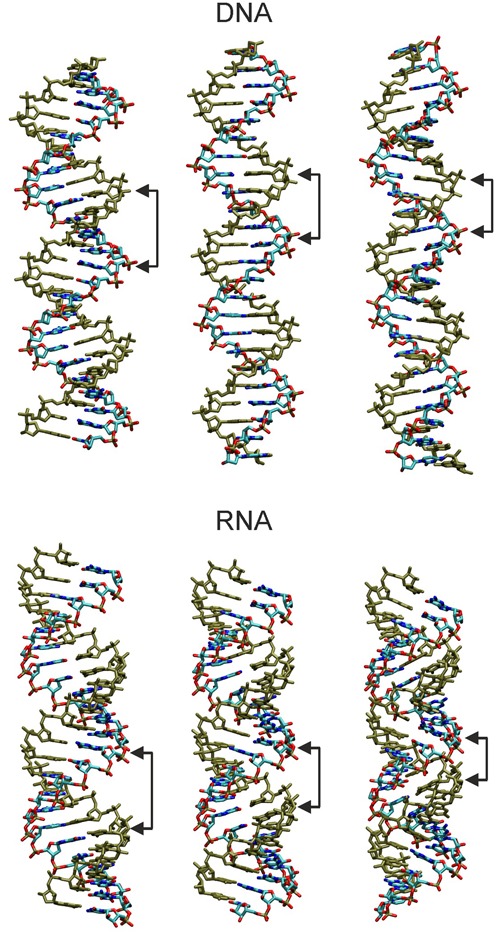
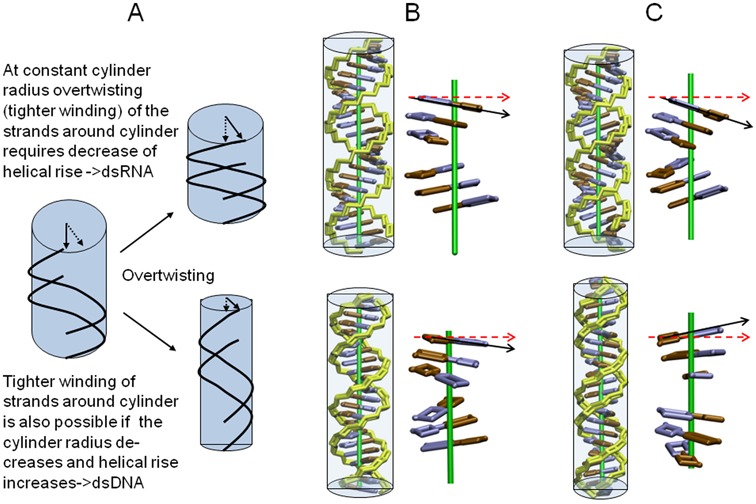
Double stranded helical DNA and RNA are flexible molecules that can undergo global conformational fluctuations. Their bending, twisting and stretching deformabilities are of similar magnitude. However, recent single-molecule experiments revealed a striking qualitative difference indicating an opposite sign for the twist-stretch couplings of dsDNA and dsRNA [Lipfert et al. 2014. Proc. Natl. Acad. Sci. U.S.A. 111, 15408] that is not explained by existing models. Employing unconstrained Molecular Dynamics (MD) simulations we are able to reproduce the qualitatively different twist-stretch coupling for dsDNA and dsRNA in semi-quantitative agreement with experiment. Similar results are also found in simulations that include an external torque to induce over- or unwinding of DNA and RNA. Detailed analysis of the helical deformations coupled to twist indicate that the interplay of helical rise, base pair inclination and displacement from the helix axis upon twist changes are responsible for the different twist-stretch correlations. Overwinding of RNA results in more compact conformations with a narrower major groove and consequently reduced helical extension. Overwinding of DNA decreases the size of the minor groove and the resulting positive base pair inclination leads to a slender and more extended helical structure.
© The Author(s) 2015. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Alexander R.W., Eargle J., Luthey-Schulten Z. Experimental and computational determination of tRNA dynamics. FEBS Lett. 2010;584:376–386. - PubMed
-
- Marko J.F., Siggia E.D. Bending and twisting elasticity of DNA. Macromolecules. 1994;27:881–998.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
