

Inhibitory Plasticity Permits the Recruitment of CA2 Pyramidal Neurons by CA3
- PMID: 26465002
- PMCID: PMC4596021
- DOI: 10.1523/ENEURO.0049-15.2015
Inhibitory Plasticity Permits the Recruitment of CA2 Pyramidal Neurons by CA3
Erratum in
-
Correction: Nasrallah et al., Inhibitory Plasticity Permits the Recruitment of CA2 Pyramidal Neurons by CA3 (eNeuro July/August 2015, 2(4) 10.1523/ENEURO.0049-15.2015 1-12 http://dx.doi.org/10.1523/ENEURO.0049-15.2015).eNeuro. 2016 Jan 11;2(6):ENEURO.0137-15.2015. doi: 10.1523/ENEURO.0137-15.2015. eCollection 2015 Nov-Dec. eNeuro. 2016. PMID: 26984103 Free PMC article.
Abstract
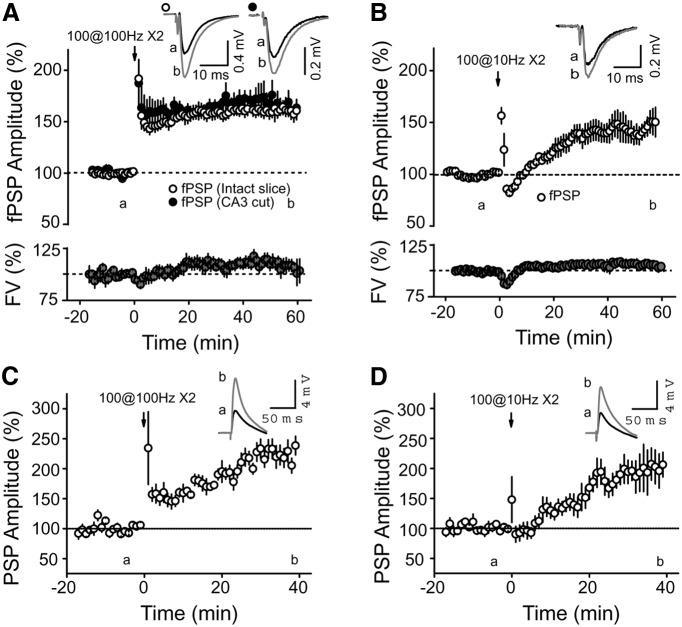
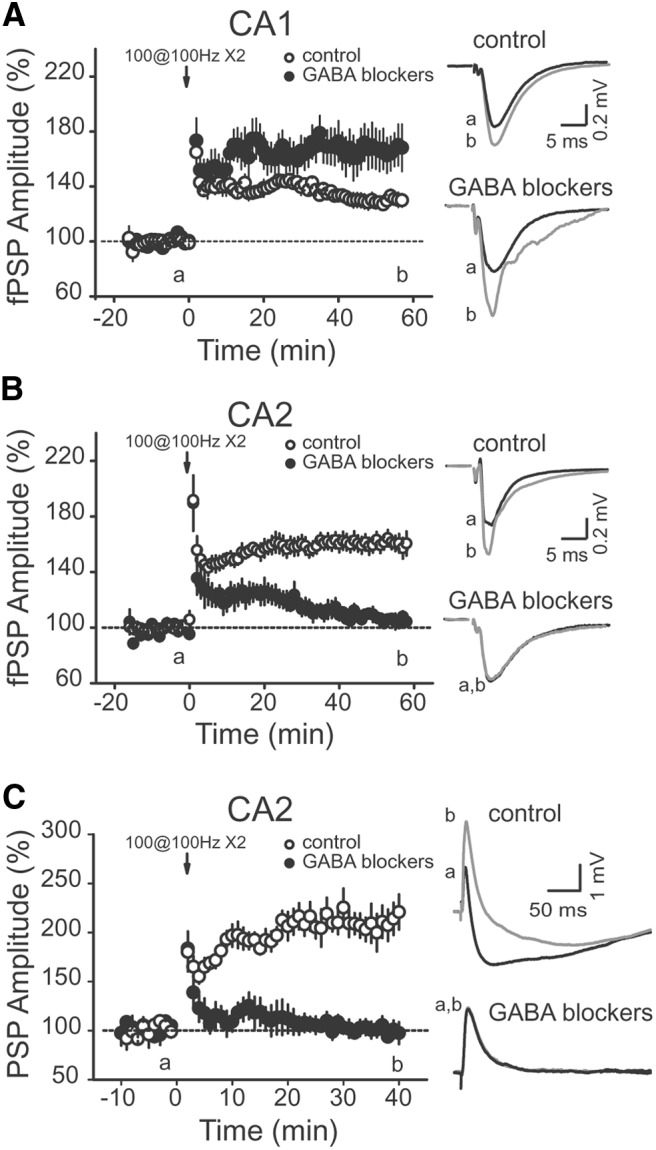
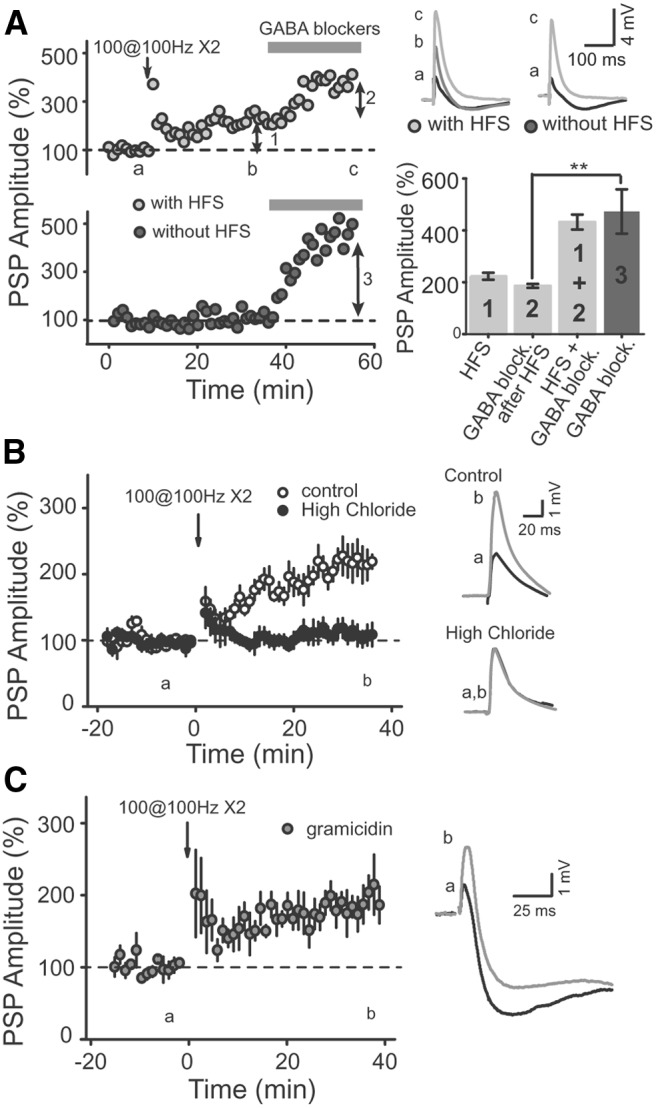
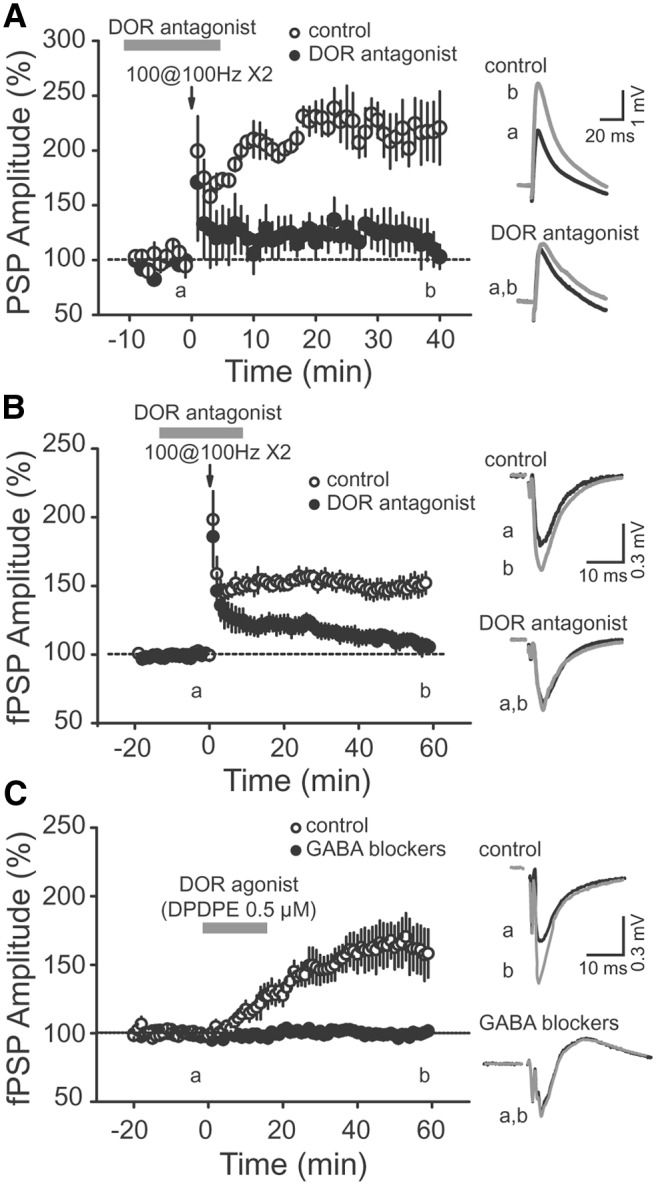
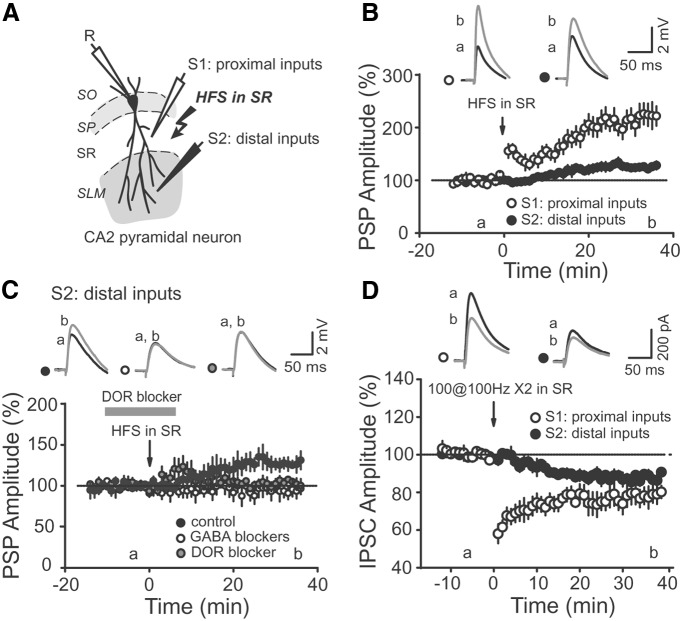
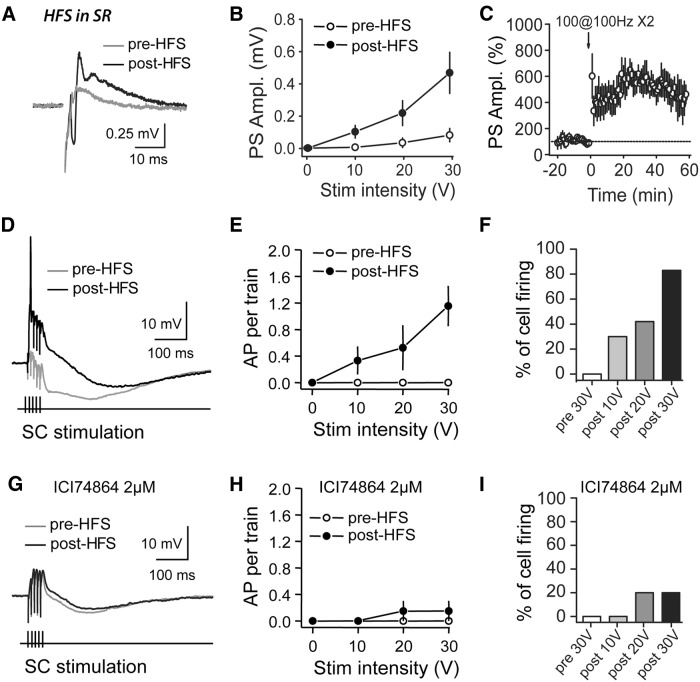
Area CA2 is emerging as an important region for hippocampal memory formation. However, how CA2 pyramidal neurons (PNs) are engaged by intrahippocampal inputs remains unclear. Excitatory transmission between CA3 and CA2 is strongly inhibited and is not plastic. We show in mice that different patterns of activity can in fact increase the excitatory drive between CA3 and CA2. We provide evidence that this effect is mediated by a long-term depression at inhibitory synapses (iLTD), as it is evoked by the same protocols and shares the same pharmacology. In addition, we show that the net excitatory drive of distal inputs is also increased after iLTD induction. The disinhibitory increase in excitatory drive is sufficient to allow CA3 inputs to evoke action potential firing in CA2 PNs. Thus, these data reveal that the output of CA2 PNs can be gated by the unique activity-dependent plasticity of inhibitory neurons in area CA2.
Keywords: area CA2; disinhibition; hippocampus; interneuron; long-term depression; δ opioid receptor.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Benes FM (1999) Evidence for altered trisynaptic circuitry in schizophrenic hippocampus. Biol Psychiatry 46:589–599. - PubMed
-
- Benes FM, Kwok EW, Vincent SL, Todtenkopf MS (1998) A reduction of nonpyramidal cells in sector CA2 of schizophrenics and manic depressives. Biol Psychiatry 44:88–97. - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous