

Absence of the Yeast Hsp31 Chaperones of the DJ-1 Superfamily Perturbs Cytoplasmic Protein Quality Control in Late Growth Phase
- PMID: 26466368
- PMCID: PMC4605529
- DOI: 10.1371/journal.pone.0140363
Absence of the Yeast Hsp31 Chaperones of the DJ-1 Superfamily Perturbs Cytoplasmic Protein Quality Control in Late Growth Phase
Abstract
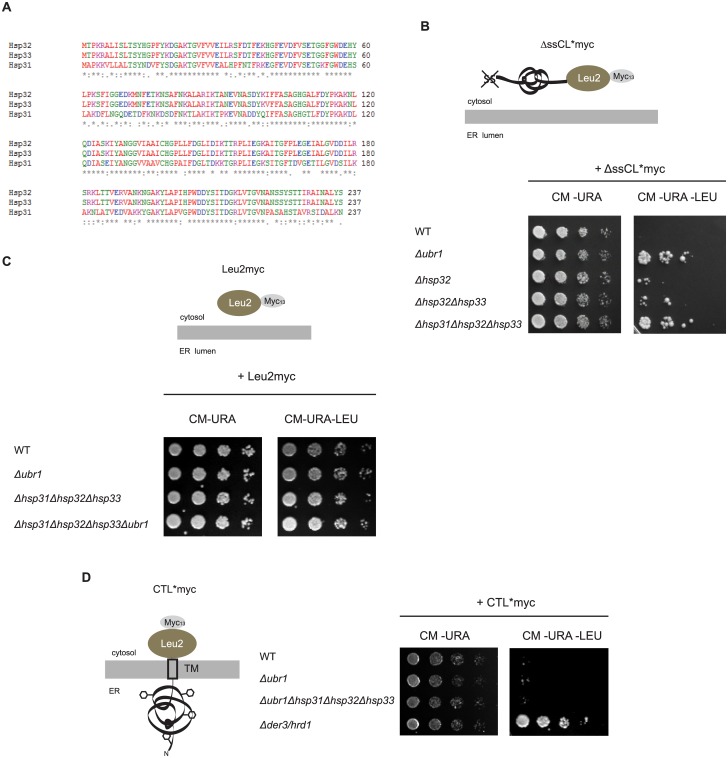
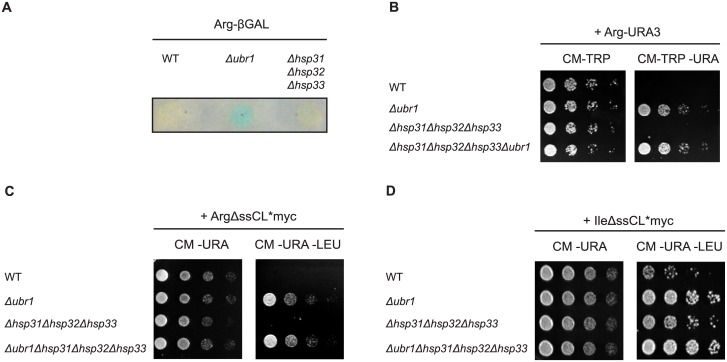
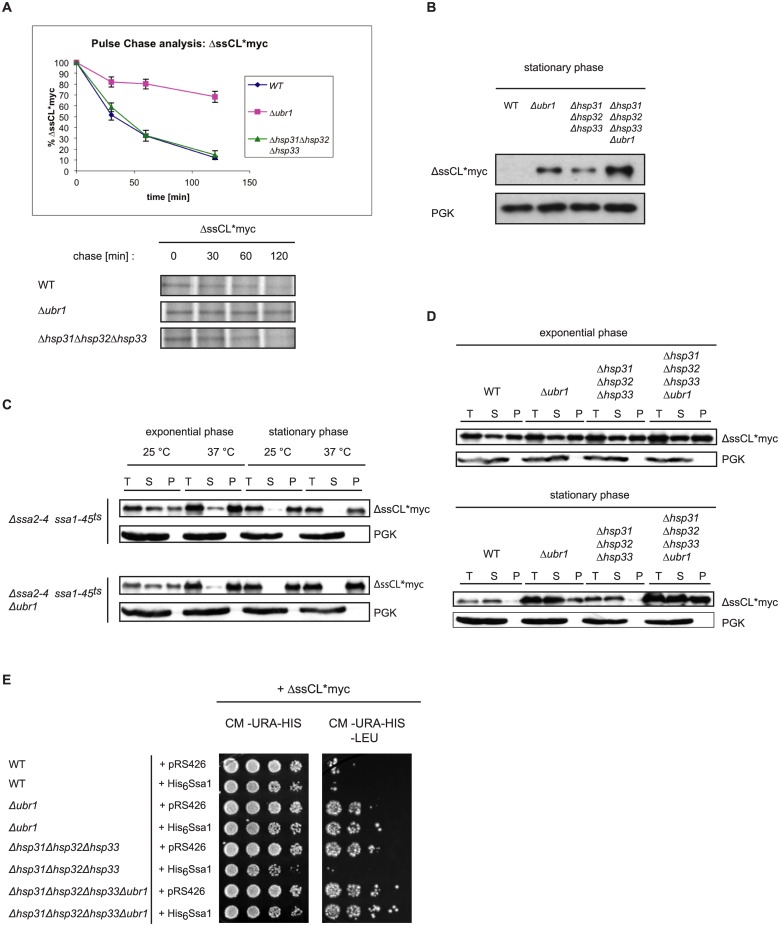
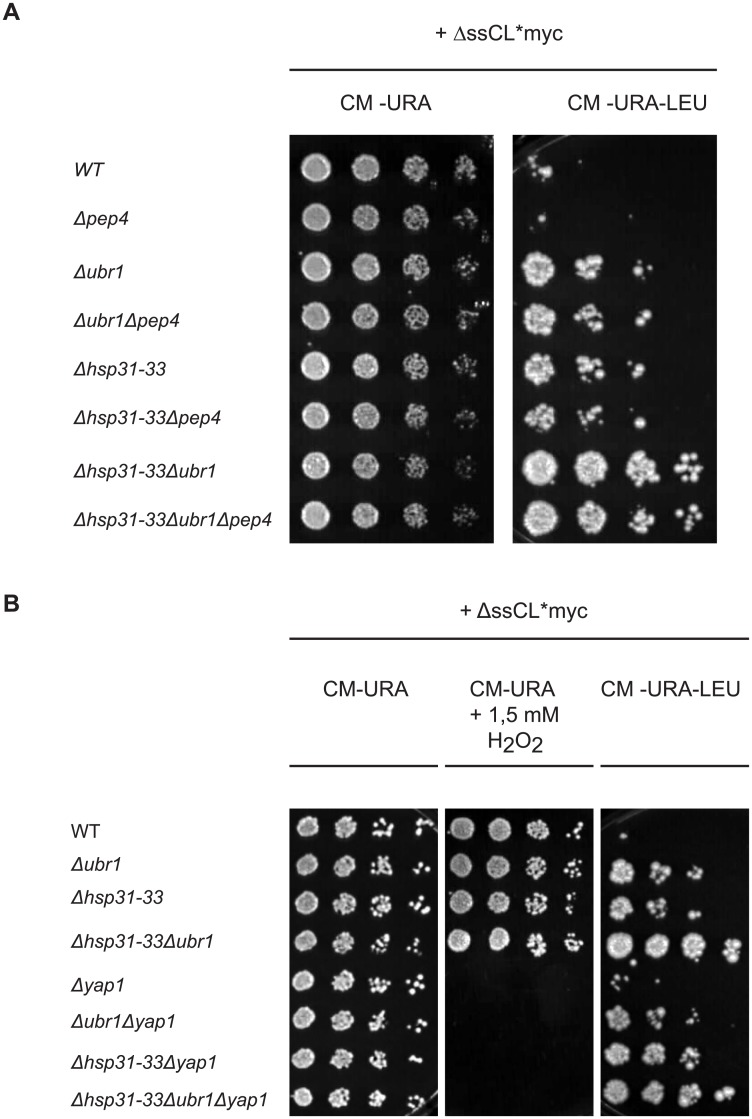
The Saccharomyces cerevisiae heat shock proteins Hsp31, Hsp32, Hsp33 and Hsp34 belong to the DJ-1/ThiJ/PfpI superfamily which includes the human protein DJ-1 (PARK7) as the most prominent member. Mutations in the DJ-1 gene are directly linked to autosomal recessive, early-onset Parkinson's disease. DJ-1 acts as an oxidative stress-induced chaperone preventing aggregation and fibrillation of α-synuclein, a critical factor in the development of the disease. In vivo assays in Saccharomyces cerevisiae using the model substrate ΔssCPY*Leu2myc (ΔssCL*myc) as an aggregation-prone misfolded cytoplasmic protein revealed an influence of the Hsp31 chaperone family on the steady state level of this substrate. In contrast to the ubiquitin ligase of the N-end rule pathway Ubr1, which is known to be prominently involved in the degradation process of misfolded cytoplasmic proteins, the absence of the Hsp31 chaperone family does not impair the degradation of newly synthesized misfolded substrate. Also degradation of substrates with strong affinity to Ubr1 like those containing the type 1 N-degron arginine is not affected by the absence of the Hsp31 chaperone family. Epistasis analysis indicates that one function of the Hsp31 chaperone family resides in a pathway overlapping with the Ubr1-dependent degradation of misfolded cytoplasmic proteins. This pathway gains relevance in late growth phase under conditions of nutrient limitation. Additionally, the Hsp31 chaperones seem to be important for maintaining the cellular Ssa Hsp70 activity which is important for Ubr1-dependent degradation.
Conflict of interest statement
Figures

References
-
- Parsell DA, Kowal AS, Singer MA, Lindquist S. Protein disaggregation mediated by heat-shock protein Hsp104. Nature. 1994;372(6505):475–8. . - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
