

Telomerase activation by genomic rearrangements in high-risk neuroblastoma
- PMID: 26466568
- PMCID: PMC4881306
- DOI: 10.1038/nature14980
Telomerase activation by genomic rearrangements in high-risk neuroblastoma
Abstract
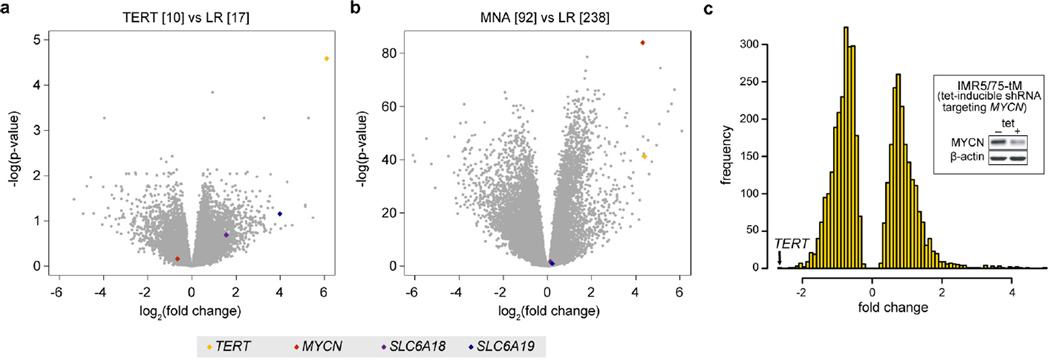
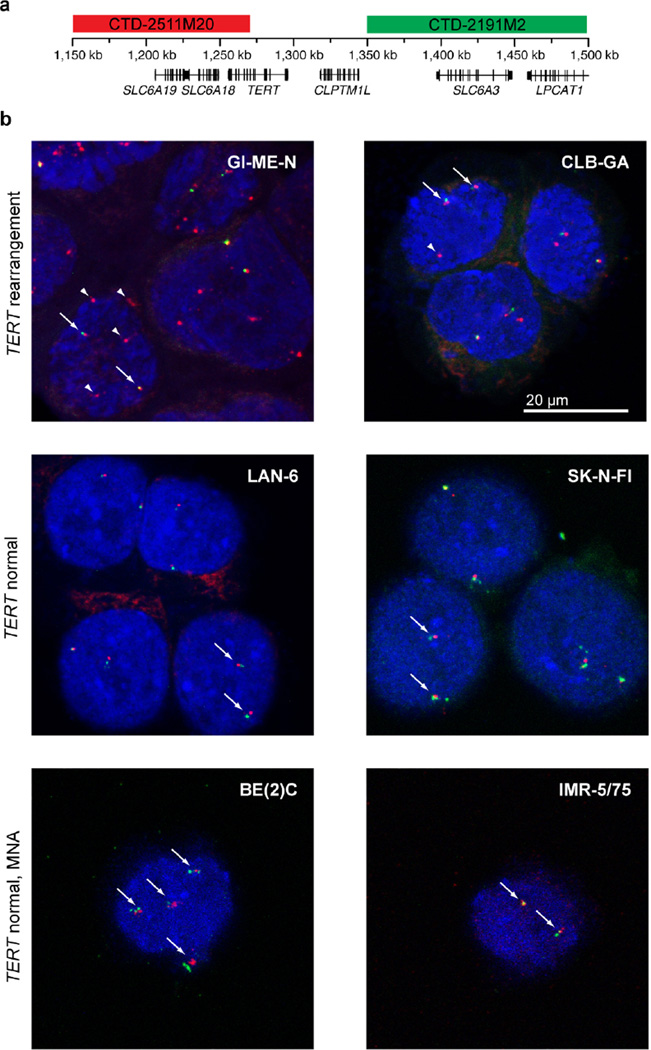
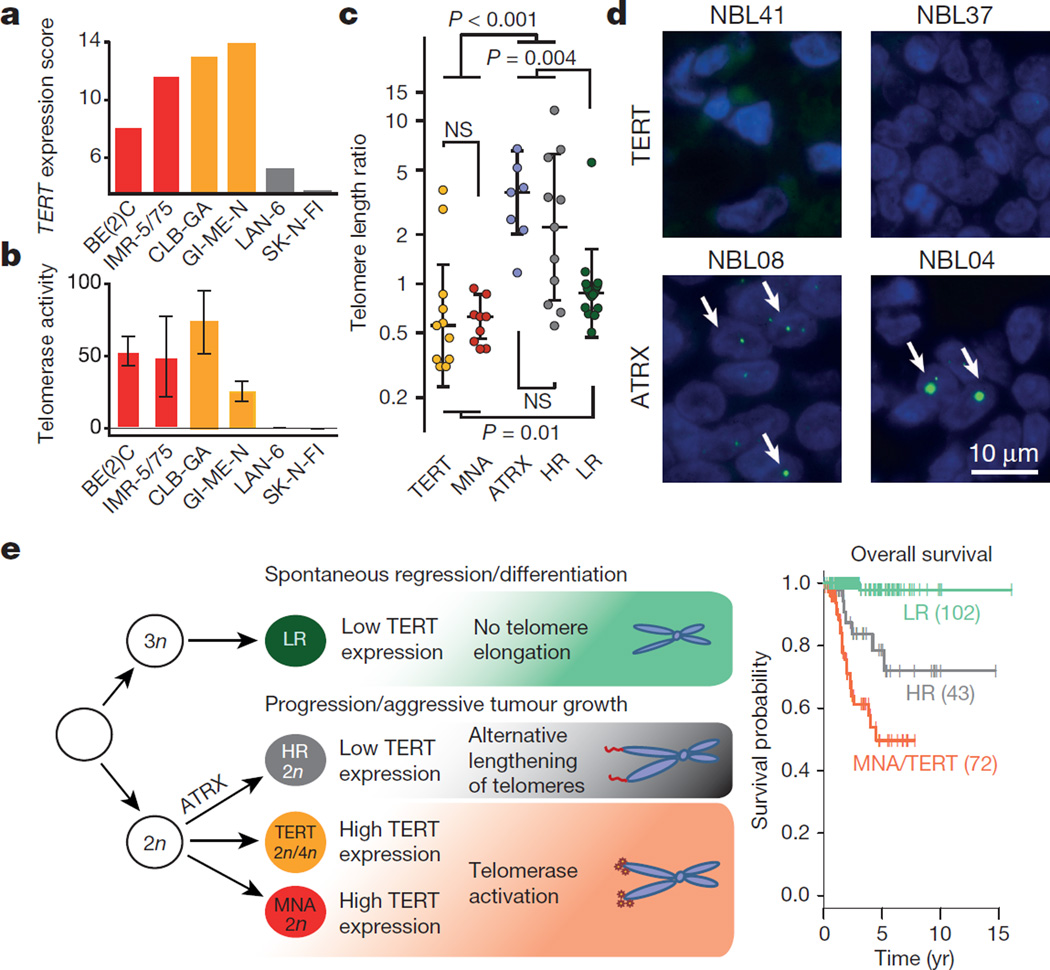
Neuroblastoma is a malignant paediatric tumour of the sympathetic nervous system. Roughly half of these tumours regress spontaneously or are cured by limited therapy. By contrast, high-risk neuroblastomas have an unfavourable clinical course despite intensive multimodal treatment, and their molecular basis has remained largely elusive. Here we have performed whole-genome sequencing of 56 neuroblastomas (high-risk, n = 39; low-risk, n = 17) and discovered recurrent genomic rearrangements affecting a chromosomal region at 5p15.33 proximal of the telomerase reverse transcriptase gene (TERT). These rearrangements occurred only in high-risk neuroblastomas (12/39, 31%) in a mutually exclusive fashion with MYCN amplifications and ATRX mutations, which are known genetic events in this tumour type. In an extended case series (n = 217), TERT rearrangements defined a subgroup of high-risk tumours with particularly poor outcome. Despite a large structural diversity of these rearrangements, they all induced massive transcriptional upregulation of TERT. In the remaining high-risk tumours, TERT expression was also elevated in MYCN-amplified tumours, whereas alternative lengthening of telomeres was present in neuroblastomas without TERT or MYCN alterations, suggesting that telomere lengthening represents a central mechanism defining this subtype. The 5p15.33 rearrangements juxtapose the TERT coding sequence to strong enhancer elements, resulting in massive chromatin remodelling and DNA methylation of the affected region. Supporting a functional role of TERT, neuroblastoma cell lines bearing rearrangements or amplified MYCN exhibited both upregulated TERT expression and enzymatic telomerase activity. In summary, our findings show that remodelling of the genomic context abrogates transcriptional silencing of TERT in high-risk neuroblastoma and places telomerase activation in the centre of transformation in a large fraction of these tumours.
Figures










References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
