

Overexpression of Cystathionine γ-Lyase Suppresses Detrimental Effects of Spinocerebellar Ataxia Type 3
- PMID: 26467707
- PMCID: PMC4749487
- DOI: 10.2119/molmed.2015.00221
Overexpression of Cystathionine γ-Lyase Suppresses Detrimental Effects of Spinocerebellar Ataxia Type 3
Abstract
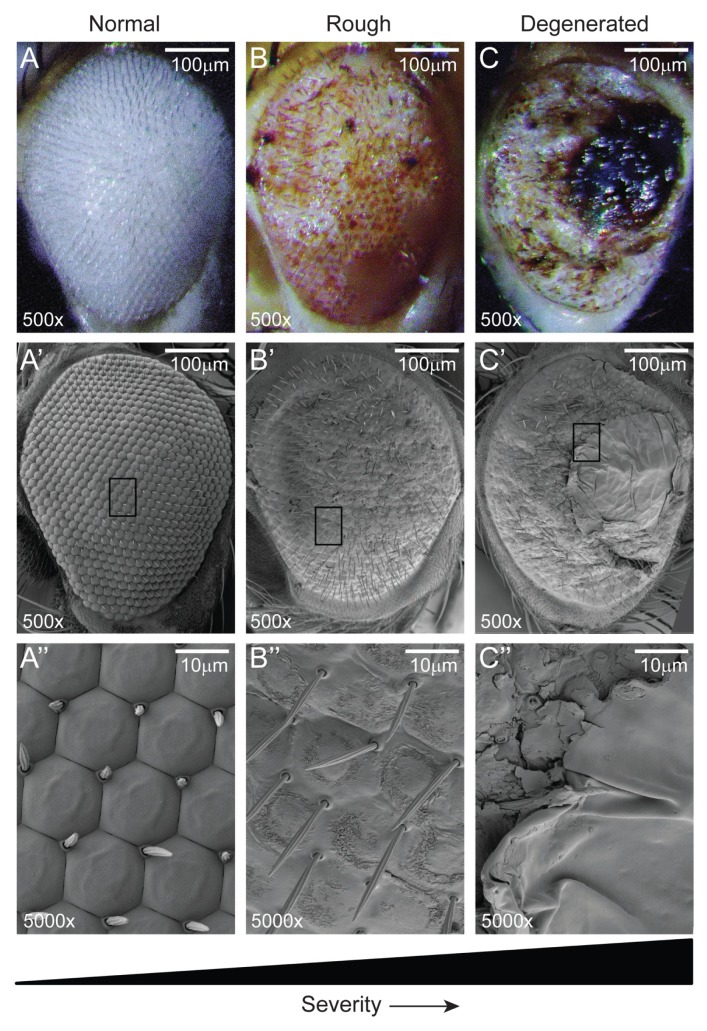
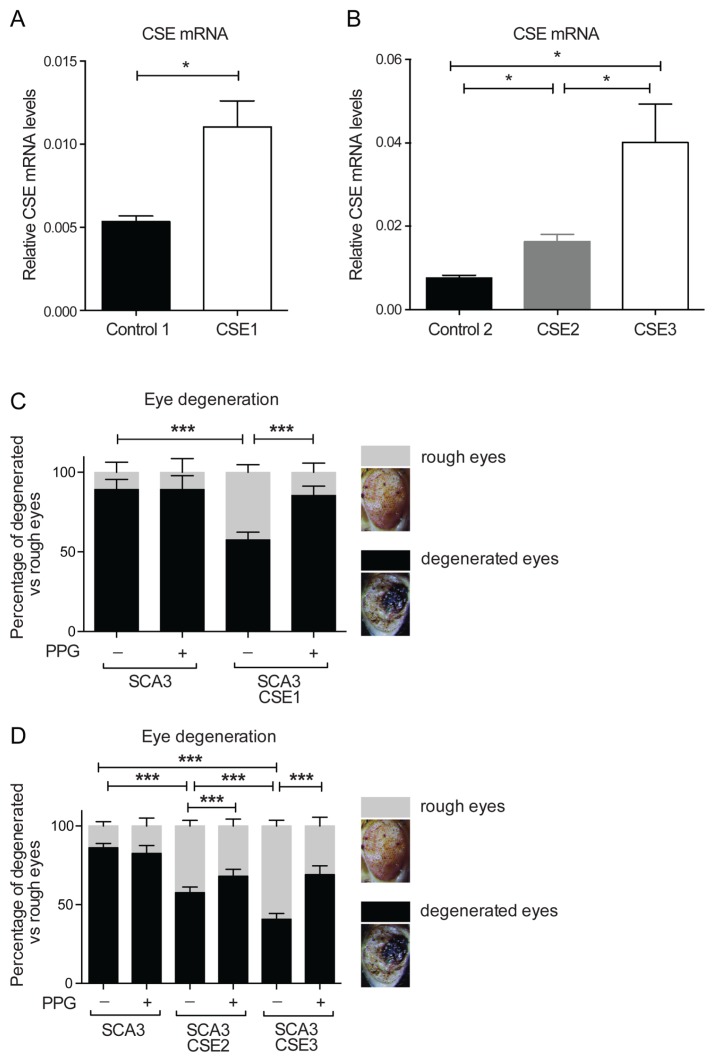
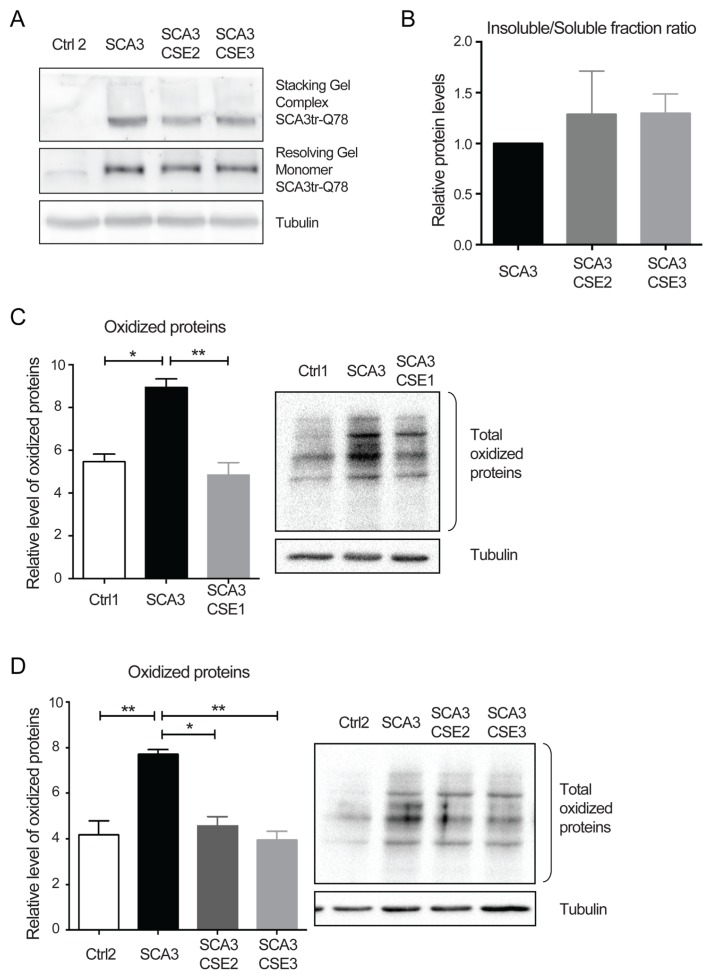
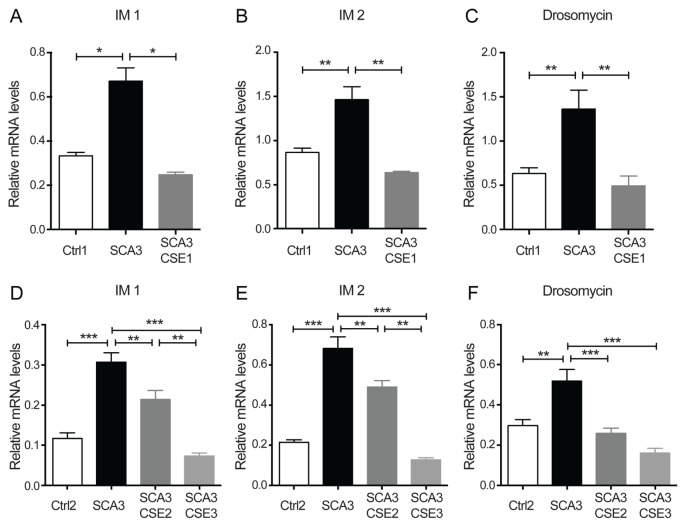
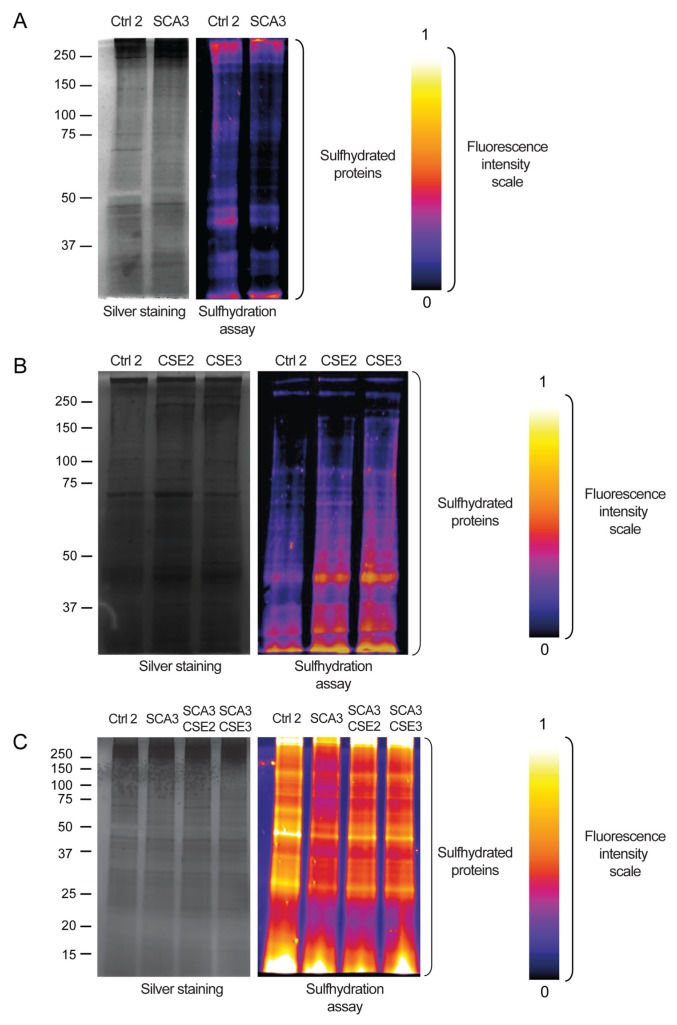
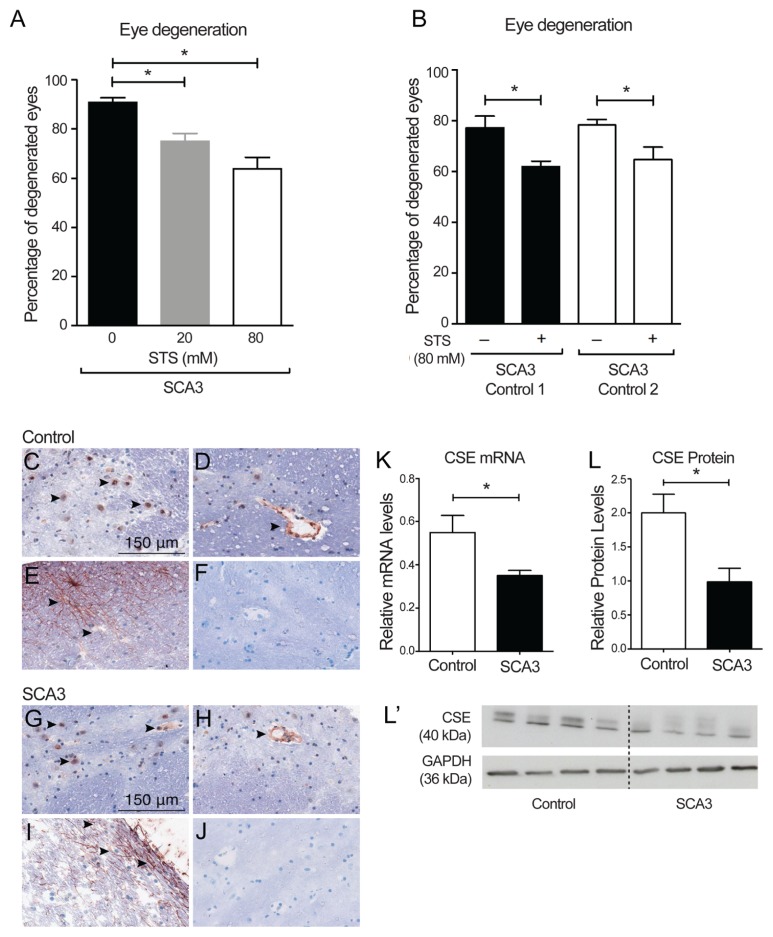
Spinocerebellar ataxia type 3 (SCA3) is a polyglutamine (polyQ) disorder caused by a CAG repeat expansion in the ataxin-3 (ATXN3) gene resulting in toxic protein aggregation. Inflammation and oxidative stress are considered secondary factors contributing to the progression of this neurodegenerative disease. There is no cure that halts or reverses the progressive neurodegeneration of SCA3. Here we show that overexpression of cystathionine γ-lyase, a central enzyme in cysteine metabolism, is protective in a Drosophila model for SCA3. SCA3 flies show eye degeneration, increased oxidative stress, insoluble protein aggregates, reduced levels of protein persulfidation and increased activation of the innate immune response. Overexpression of Drosophila cystathionine γ-lyase restores protein persulfidation, decreases oxidative stress, dampens the immune response and improves SCA3-associated tissue degeneration. Levels of insoluble protein aggregates are not altered; therefore, the data implicate a modifying role of cystathionine γ-lyase in ameliorating the downstream consequence of protein aggregation leading to protection against SCA3-induced tissue degeneration. The cystathionine γ-lyase expression is decreased in affected brain tissue of SCA3 patients, suggesting that enhancers of cystathionine γ-lyase expression or activity are attractive candidates for future therapies.
Figures
References
-
- Zoghbi HY, Orr HT. Glutamine repeats and neurodegeneration. Annu Rev Neurosci. 2000;23:217–47. - PubMed
-
- Hageman J, et al. A DNAJB chaperone subfamily with HDAC-dependent activities suppresses toxic protein aggregation. Mol Cell. 2010;37:355–69. - PubMed
-
- Dueñas AM, Goold R, Giunti P. Molecular pathogenesis of spinocerebellar ataxias. Brain. 2006;129:1357–70. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
