

Comparative Efficacy of Monoclonal Antibodies That Bind to Different Epitopes of the 2009 Pandemic H1N1 Influenza Virus Neuraminidase
- PMID: 26468531
- PMCID: PMC4702561
- DOI: 10.1128/JVI.01756-15
Comparative Efficacy of Monoclonal Antibodies That Bind to Different Epitopes of the 2009 Pandemic H1N1 Influenza Virus Neuraminidase
Abstract
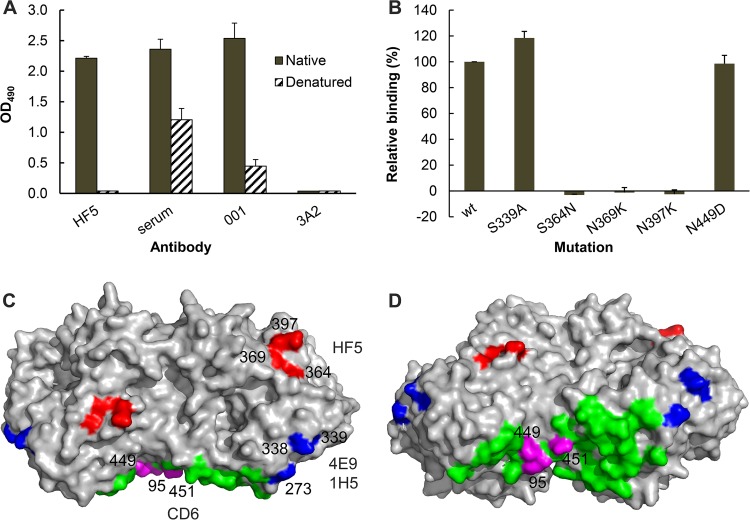
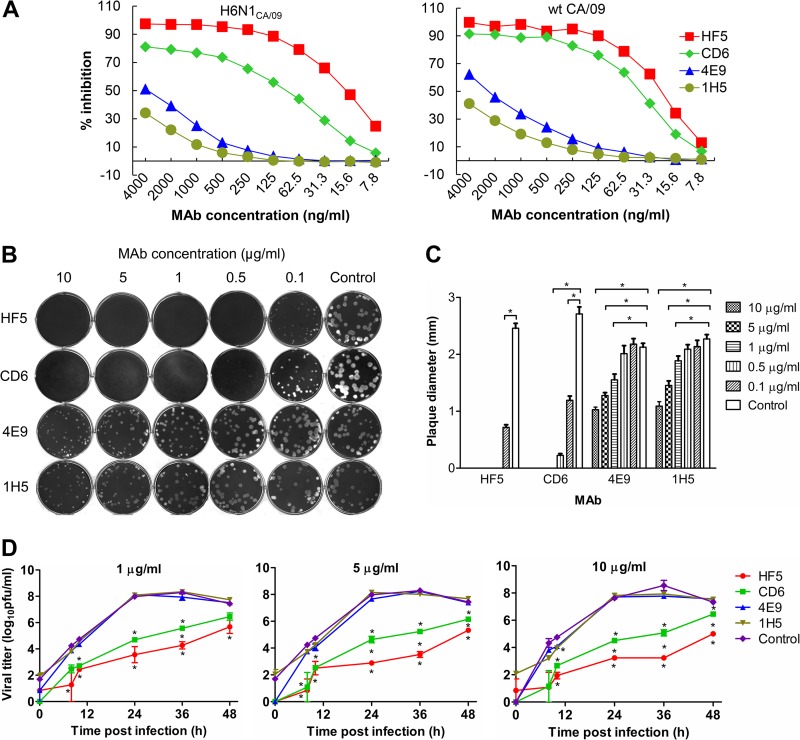
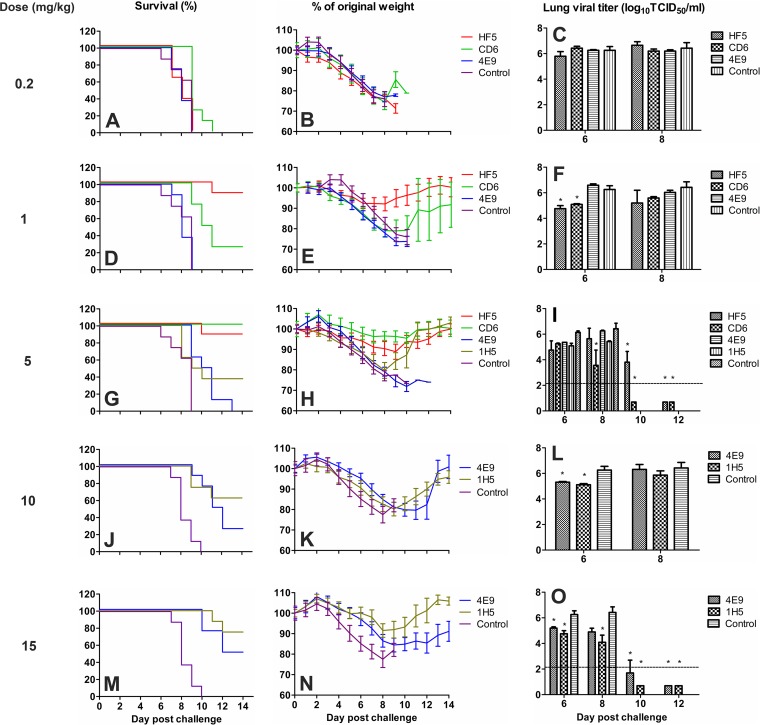
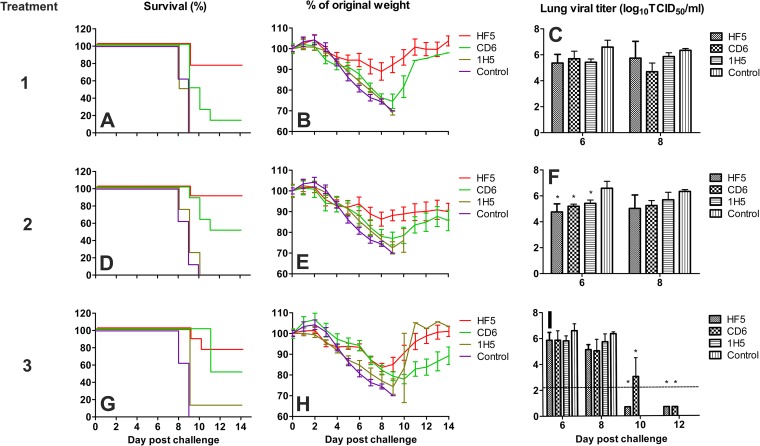
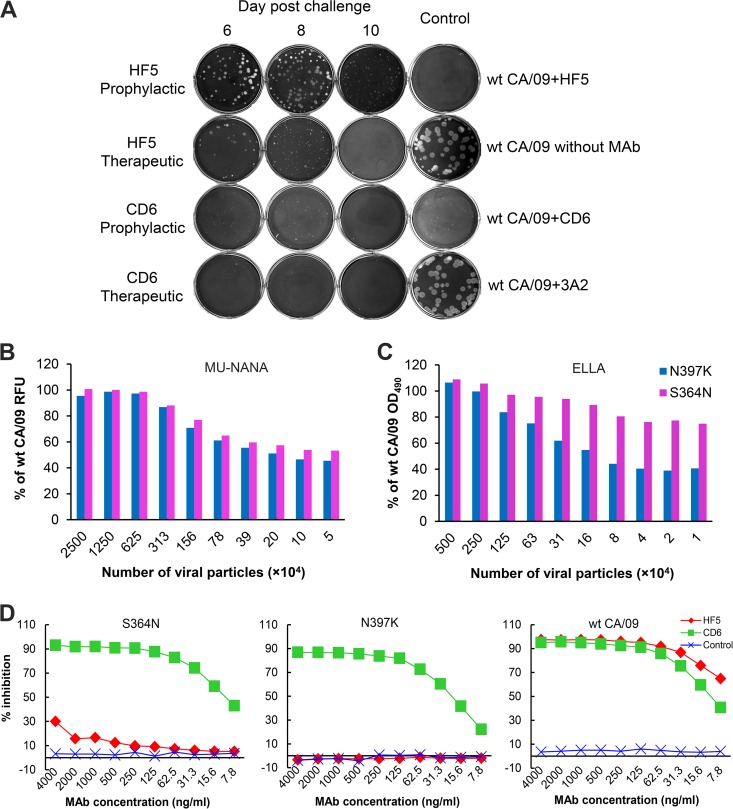
Antibodies against the neuraminidase (NA) of influenza virus correlate with resistance against disease, but the effectiveness of antibodies against different NA epitopes has not been compared. In the present study, we evaluated the in vitro and in vivo efficacies of four monoclonal antibodies (MAbs): HF5 and CD6, which are specific to two different epitopes in the NA of 2009 pandemic H1N1 (pH1N1) virus, and 4E9 and 1H5, which are specific to a conserved epitope in the NA of both H1N1 and H5N1 viruses. In the in vitro assays, HF5 and CD6 inhibited virus spread and growth more effectively than 4E9 and 1H5, with HF5 being the most effective inhibitor. When administered prophylactically at 5 mg/kg of body weight, HF5 and CD6 protected ~90 to 100% of DBA/2 mice against lethal wild-type pH1N1 virus challenge; however, at a lower dose (1 mg/kg), HF5 protected ~90% of mice, whereas CD6 protected only 25% of mice. 4E9 and 1H5 were less effective than HF5 and CD6, as indicated by the partial protection achieved even at doses as high as 15 mg/kg. When administered therapeutically, HF5 protected a greater proportion of mice against lethal pH1N1 challenge than CD6. However, HF5 quickly selected pH1N1 virus escape mutants in both prophylactic and therapeutic treatments, while CD6 did not. Our findings confirm the important role of NA-specific antibodies in immunity to influenza virus and provide insight into the properties of NA antibodies that may serve as good candidates for therapeutics against influenza.
Importance: Neuraminidase (NA) is one of the major surface proteins of influenza virus, serving as an important target for antivirals and therapeutic antibodies. The impact of NA-specific antibodies on NA activity and virus replication is likely to depend on where the antibody binds. Using in vitro assays and the mouse model, we compared the inhibitory/protective efficacy of four mouse monoclonal antibodies (MAbs) that bind to different sites within the 2009 pandemic H1N1 (pH1N1) virus NA. The ability of each MAb to protect mice against lethal pH1N1 infection corresponded to its ability to inhibit NA activity in vitro; however, the MAb that was the most effective inhibitor of NA activity selected pH1N1 escape variants in vivo. One of the tested MAbs, which binds to a conserved region in the NA of pH1N1 virus, inhibited NA activity but did not result in escape variants, highlighting its suitability for development as a therapeutic agent.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
