

Conformations of tissue plasminogen activator (tPA) orchestrate neuronal survival by a crosstalk between EGFR and NMDAR
- PMID: 26469972
- PMCID: PMC4632317
- DOI: 10.1038/cddis.2015.296
Conformations of tissue plasminogen activator (tPA) orchestrate neuronal survival by a crosstalk between EGFR and NMDAR
Abstract
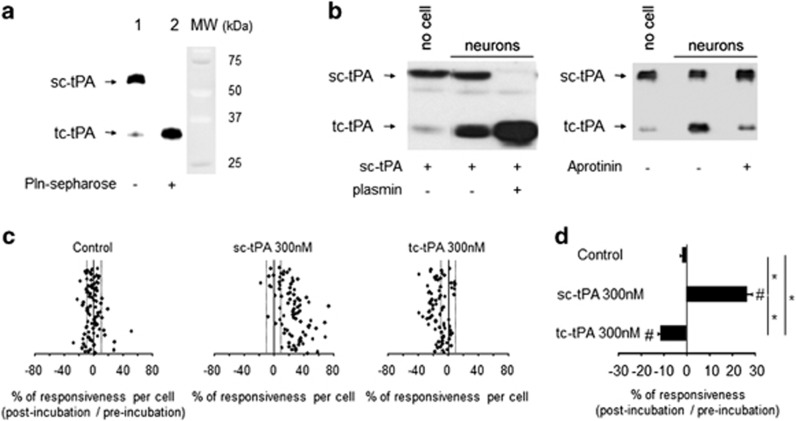
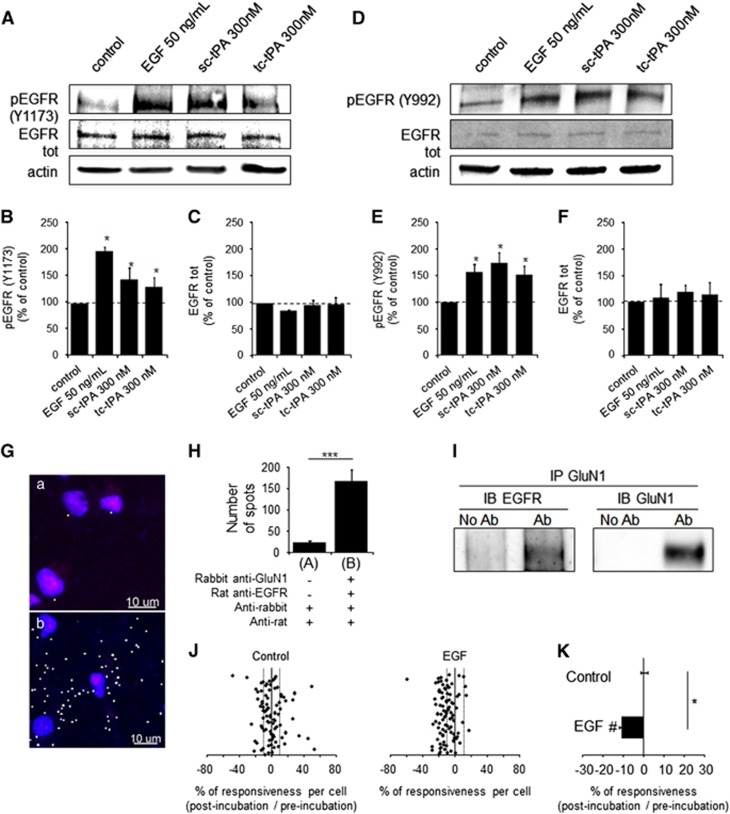
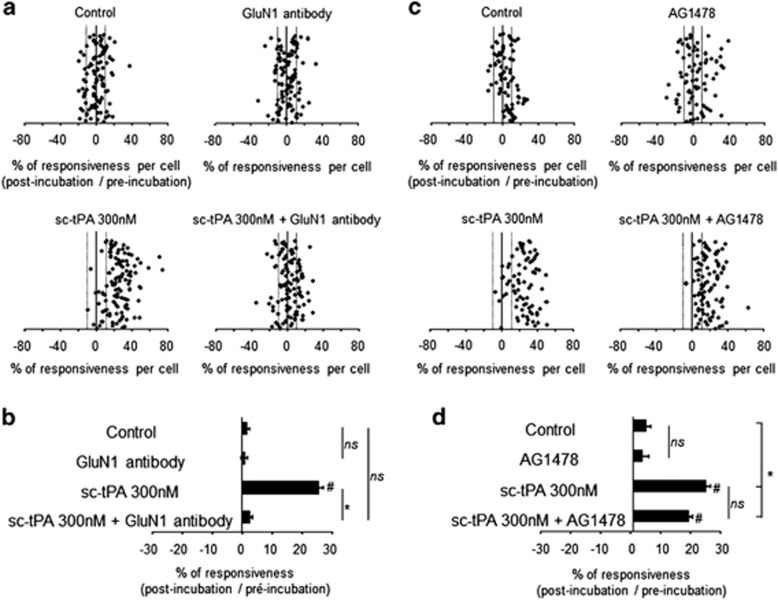
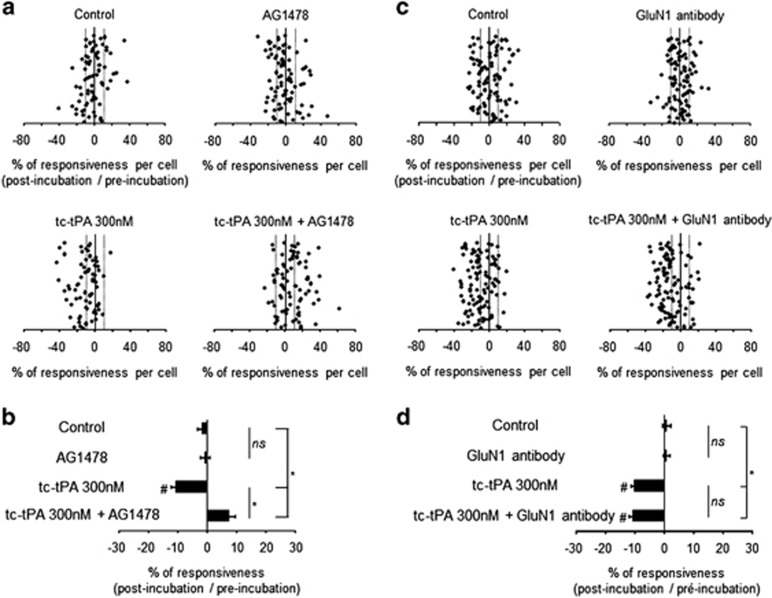
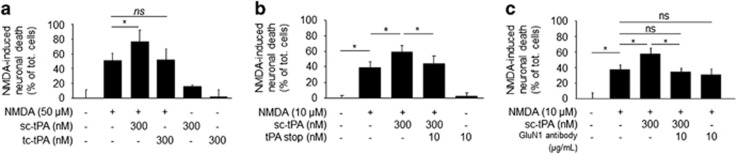
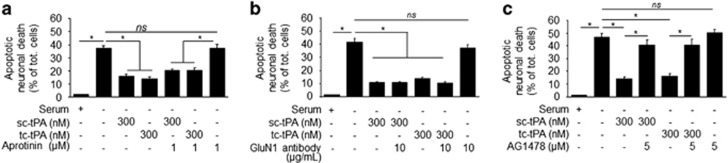
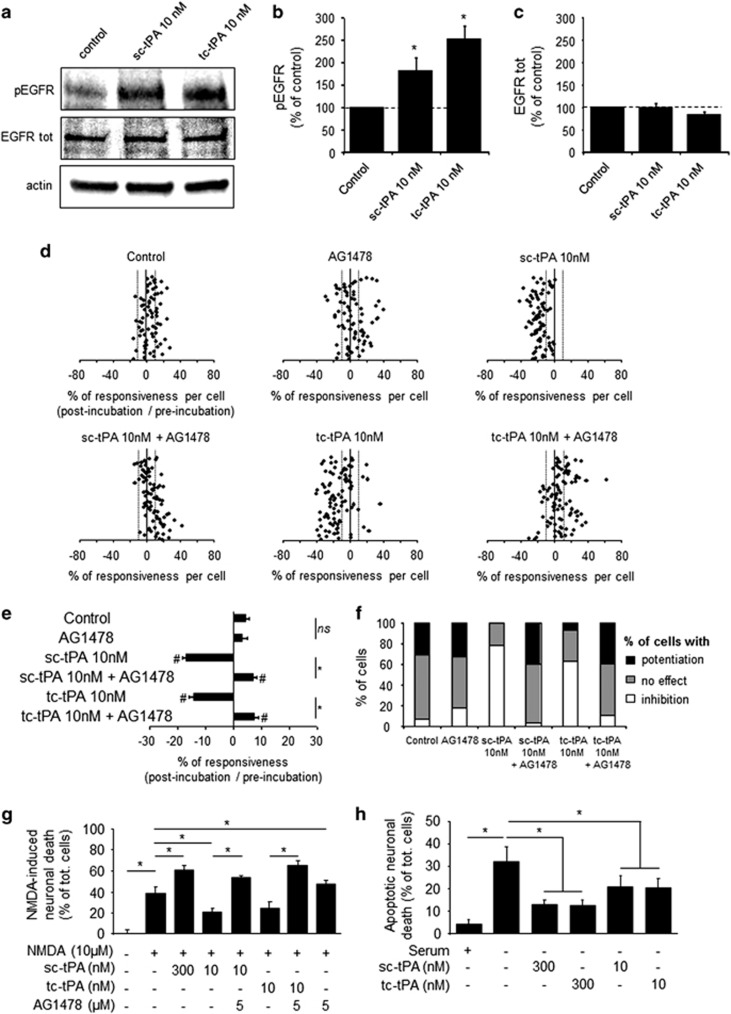
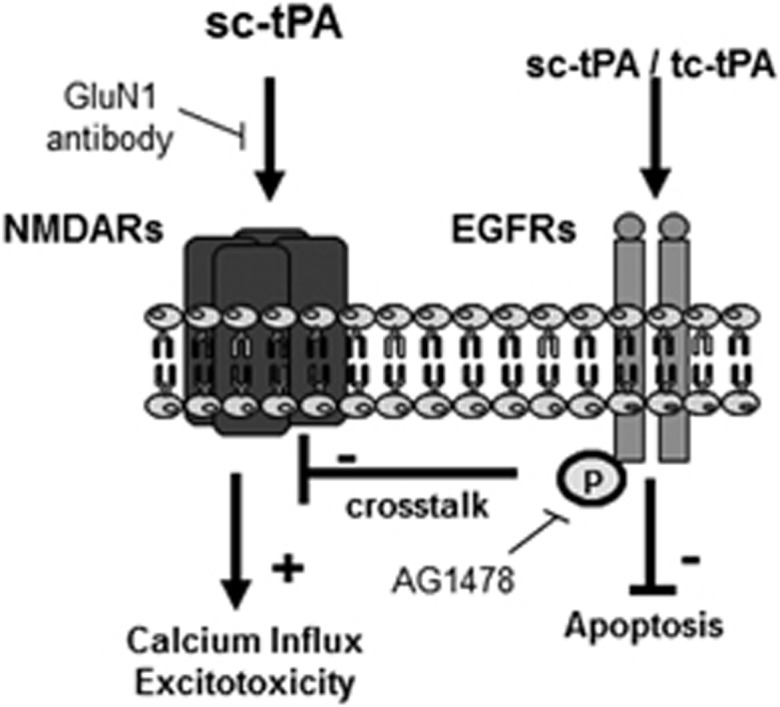
Tissue-type plasminogen activator (tPA) is a pleiotropic serine protease of the central nervous system (CNS) with reported neurotrophic and neurotoxic functions. Produced and released under its single chain form (sc), the sc-tPA can be cleaved by plasmin or kallikrein in a two chain form, tc-tPA. Although both sc-tPA and tc-tPA display a similar fibrinolytic activity, we postulated here that these two conformations of tPA (sc-tPA and tc-tPA) could differentially control the effects of tPA on neuronal survival. Using primary cultures of mouse cortical neurons, our present study reveals that sc-tPA is the only one capable to promote N-methyl-D-aspartate receptor (NMDAR)-induced calcium influx and subsequent excitotoxicity. In contrast, both sc-tPA and tc-tPA are capable to activate epidermal growth factor receptors (EGFRs), a mechanism mediating the antiapoptotic effects of tPA. Interestingly, we revealed a tPA dependent crosstalk between EGFR and NMDAR in which a tPA-dependent activation of EGFRs leads to downregulation of NMDAR signaling and to subsequent neurotrophic effects.
Figures
References
-
- 1Thorsen S, Glas-Greenwalt P, Astrup T. Differences in the binding to fibrin of urokinase and tissue plasminogen activator. Thromb Diath Haemorrh 1972; 28: 65–74. - PubMed
-
- 5Gorter JA, Van Vliet EA, Rauwerda H, Breit T, Stad R, van Schaik L et al. Dynamic changes of proteases and protease inhibitors revealed by microarray analysis in CA3 and entorhinal cortex during epileptogenesis in the rat. Epilepsia 2007; 48: 53–64. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
