

Codon reassignment to facilitate genetic engineering and biocontainment in the chloroplast of Chlamydomonas reinhardtii
- PMID: 26471875
- PMCID: PMC5102678
- DOI: 10.1111/pbi.12490
Codon reassignment to facilitate genetic engineering and biocontainment in the chloroplast of Chlamydomonas reinhardtii
Abstract
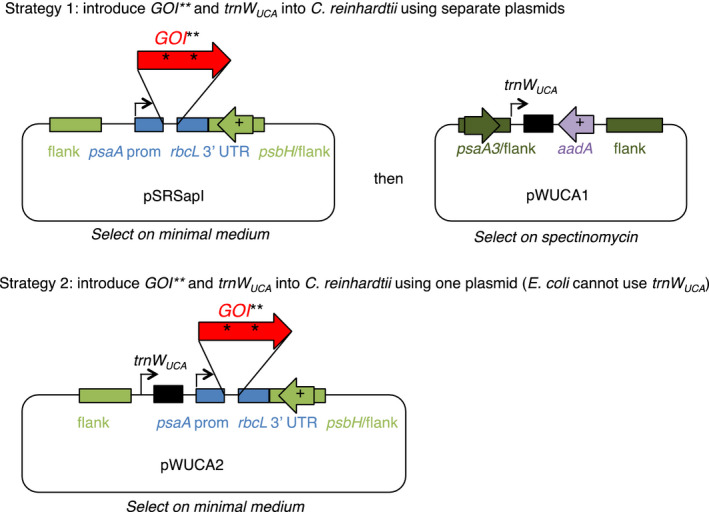
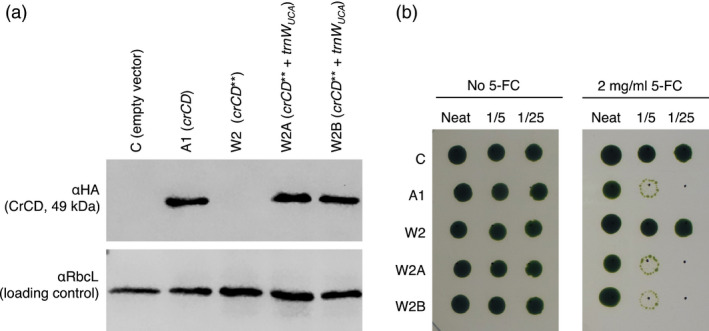
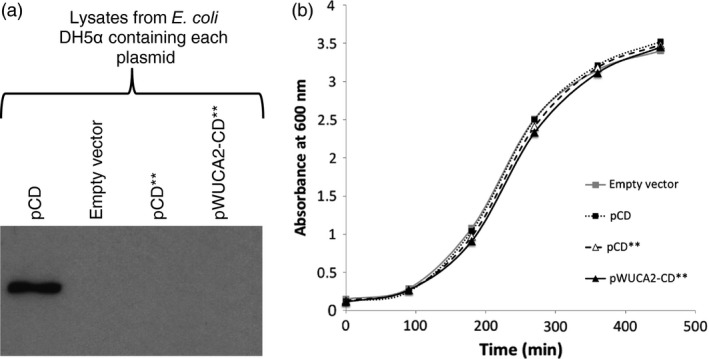
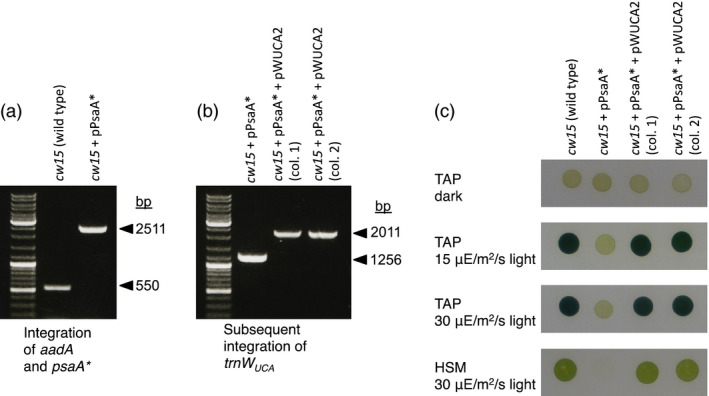
There is a growing interest in the use of microalgae as low-cost hosts for the synthesis of recombinant products such as therapeutic proteins and bioactive metabolites. In particular, the chloroplast, with its small, genetically tractable genome (plastome) and elaborate metabolism, represents an attractive platform for genetic engineering. In Chlamydomonas reinhardtii, none of the 69 protein-coding genes in the plastome uses the stop codon UGA, therefore this spare codon can be exploited as a useful synthetic biology tool. Here, we report the assignment of the codon to one for tryptophan and show that this can be used as an effective strategy for addressing a key problem in chloroplast engineering: namely, the assembly of expression cassettes in Escherichia coli when the gene product is toxic to the bacterium. This problem arises because the prokaryotic nature of chloroplast promoters and ribosome-binding sites used in such cassettes often results in transgene expression in E. coli, and is a potential issue when cloning genes for metabolic enzymes, antibacterial proteins and integral membrane proteins. We show that replacement of tryptophan codons with the spare codon (UGG→UGA) within a transgene prevents functional expression in E. coli and in the chloroplast, and that co-introduction of a plastidial trnW gene carrying a modified anticodon restores function only in the latter by allowing UGA readthrough. We demonstrate the utility of this system by expressing two genes known to be highly toxic to E. coli and discuss its value in providing an enhanced level of biocontainment for transplastomic microalgae.
Keywords: Chlamydomonas reinhardtii; chloroplast; microalgae; non-sense suppression; transfer RNA.
© 2015 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd.
Figures
Similar articles
-
CITRIC: cold-inducible translational readthrough in the chloroplast of Chlamydomonas reinhardtii using a novel temperature-sensitive transfer RNA.Microb Cell Fact. 2018 Nov 24;17(1):186. doi: 10.1186/s12934-018-1033-5. Microb Cell Fact. 2018. PMID: 30474564 Free PMC article.
-
A Simple Technology for Generating Marker-Free Chloroplast Transformants of the Green Alga Chlamydomonas reinhardtii.Methods Mol Biol. 2021;2317:293-304. doi: 10.1007/978-1-0716-1472-3_17. Methods Mol Biol. 2021. PMID: 34028777
-
Multigenic engineering of the chloroplast genome in the green alga Chlamydomonas reinhardtii.Microbiology (Reading). 2020 Jun;166(6):510-515. doi: 10.1099/mic.0.000910. Microbiology (Reading). 2020. PMID: 32250732 Free PMC article.
-
Outlook in the application of Chlamydomonas reinhardtii chloroplast as a platform for recombinant protein production.Biotechnol Genet Eng Rev. 2016 Apr-Oct;32(1-2):92-106. doi: 10.1080/02648725.2017.1307673. Epub 2017 Mar 30. Biotechnol Genet Eng Rev. 2016. PMID: 28359189 Review.
-
Mechanisms governing codon usage bias and the implications for protein expression in the chloroplast of Chlamydomonas reinhardtii.Plant J. 2022 Nov;112(4):919-945. doi: 10.1111/tpj.15970. Epub 2022 Oct 19. Plant J. 2022. PMID: 36071273 Free PMC article. Review.
Cited by
-
Microalgae for bioremediation: advances, challenges, and public perception on genetic engineering.BMC Plant Biol. 2024 Dec 27;24(1):1261. doi: 10.1186/s12870-024-05995-5. BMC Plant Biol. 2024. PMID: 39731038 Free PMC article. Review.
-
A microalgal-based preparation with synergistic cellulolytic and detoxifying action towards chemical-treated lignocellulose.Plant Biotechnol J. 2021 Jan;19(1):124-137. doi: 10.1111/pbi.13447. Epub 2020 Sep 2. Plant Biotechnol J. 2021. PMID: 32649019 Free PMC article.
-
Genetic Engineering: A Promising Tool to Engender Physiological, Biochemical, and Molecular Stress Resilience in Green Microalgae.Front Plant Sci. 2016 Mar 31;7:400. doi: 10.3389/fpls.2016.00400. eCollection 2016. Front Plant Sci. 2016. PMID: 27066043 Free PMC article. Review.
-
Selectable Markers and Reporter Genes for Engineering the Chloroplast of Chlamydomonas reinhardtii.Biology (Basel). 2018 Oct 10;7(4):46. doi: 10.3390/biology7040046. Biology (Basel). 2018. PMID: 30309004 Free PMC article. Review.
-
Use of the ptxD gene as a portable selectable marker for chloroplast transformation in Chlamydomonas reinhardtii.Mol Biotechnol. 2019 Jun;61(6):461-468. doi: 10.1007/s12033-019-00177-3. Mol Biotechnol. 2019. PMID: 30997667
References
-
- Agris, P.F. , Vendeix, F.A. and Graham, W.D. (2007) tRNA's wobble decoding of the genome: 40 years of modification. J. Mol. Biol. 366, 1–13. - PubMed
-
- Bock, R. (2015) Engineering plastid genomes: methods, tools, and applications in basic research and biotechnology. Annu. Rev. Plant Biol. 66, 211–241. - PubMed
-
- Boudreaux, B. , MacMillan, F. , Teutloff, C. , Agalarov, R. , Gu, F. , Grimaldi, S. , Bittl, R. et al. (2001) Mutations in both sides of the photosystem I reaction center identify the phylloquinone observed by electron paramagnetic resonance spectroscopy. J. Biol. Chem. 276, 37299–37306. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
