

Reduced naïve CD8(+) T-cell priming efficacy in elderly adults
- PMID: 26472076
- PMCID: PMC4717282
- DOI: 10.1111/acel.12384
Reduced naïve CD8(+) T-cell priming efficacy in elderly adults
Abstract
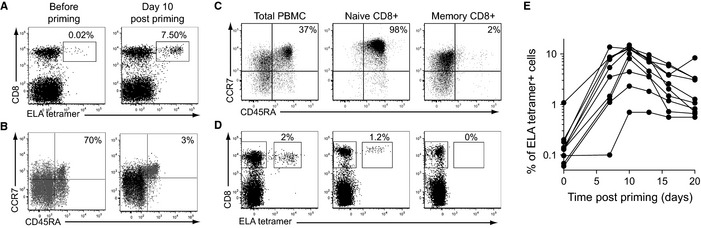
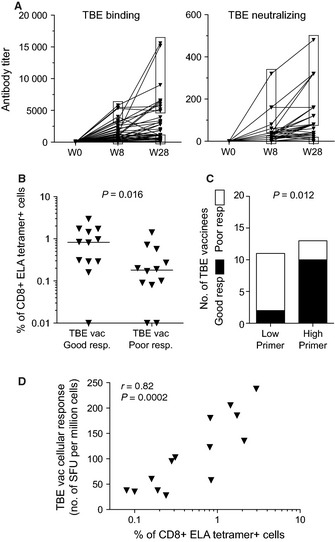
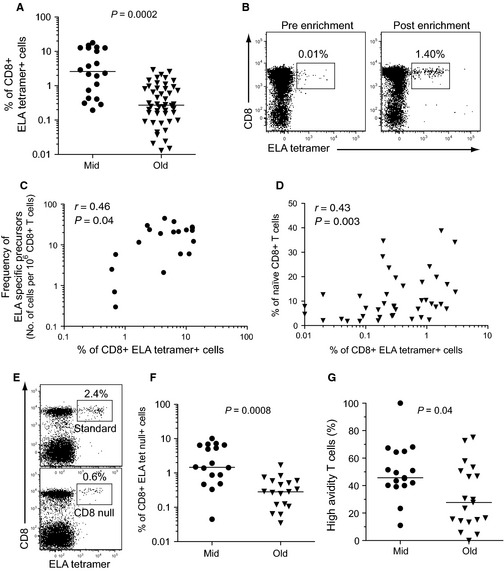
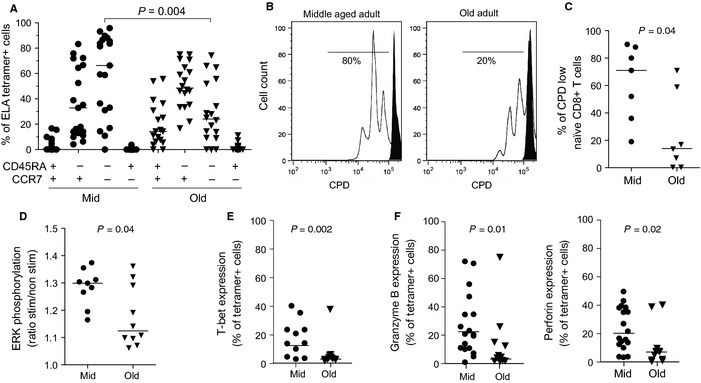
Aging is associated with impaired vaccine efficacy and increased susceptibility to infectious and malignant diseases. CD8(+) T-cells are key players in the immune response against pathogens and tumors. In aged mice, the dwindling naïve CD8(+) T-cell compartment is thought to compromise the induction of de novo immune responses, but no experimental evidence is yet available in humans. Here, we used an original in vitro assay based on an accelerated dendritic cell coculture system in unfractioned peripheral blood mononuclear cells to examine CD8(+) T-cell priming efficacy in human volunteers. Using this approach, we report that old individuals consistently mount quantitatively and qualitatively impaired de novo CD8(+) T-cell responses specific for a model antigen. Reduced CD8(+) T-cell priming capacity in vitro was further associated with poor primary immune responsiveness in vivo. This immune deficit likely arises as a consequence of intrinsic cellular defects and a reduction in the size of the naïve CD8(+) T-cell pool. Collectively, these findings provide new insights into the cellular immune insufficiencies that accompany human aging.
Keywords: aging; naïve CD8+ T-cells; priming.
© 2015 The Authors. Aging Cell published by the Anatomical Society and John Wiley & Sons Ltd.
Figures
Similar articles
-
Loss of naive T cells and repertoire constriction predict poor response to vaccination in old primates.J Immunol. 2010 Jun 15;184(12):6739-45. doi: 10.4049/jimmunol.0904193. Epub 2010 May 17. J Immunol. 2010. PMID: 20483749 Free PMC article.
-
Low Thymic Activity and Dendritic Cell Numbers Are Associated with the Immune Response to Primary Viral Infection in Elderly Humans.J Immunol. 2015 Nov 15;195(10):4699-711. doi: 10.4049/jimmunol.1500598. Epub 2015 Oct 12. J Immunol. 2015. PMID: 26459351
-
Impaired T-cell priming and proliferation in cats infected with feline immunodeficiency virus.AIDS. 1992 Mar;6(3):287-93. doi: 10.1097/00002030-199203000-00005. AIDS. 1992. PMID: 1348945
-
CD8+ T-cell priming regulated by cytokines of the innate immune system.Trends Mol Med. 2004 Aug;10(8):366-71. doi: 10.1016/j.molmed.2004.06.003. Trends Mol Med. 2004. PMID: 15310456 Review.
-
CD8+ T cell differentiation in the aging immune system: until the last clone standing.Curr Opin Immunol. 2011 Aug;23(4):549-54. doi: 10.1016/j.coi.2011.05.002. Epub 2011 Jun 12. Curr Opin Immunol. 2011. PMID: 21664807 Review.
Cited by
-
Effects of the age of vaccination on the humoral responses to a human papillomavirus vaccine.NPJ Vaccines. 2022 Mar 15;7(1):37. doi: 10.1038/s41541-022-00458-0. NPJ Vaccines. 2022. PMID: 35292655 Free PMC article.
-
Naïve CD8+ T-Cells Engage a Versatile Metabolic Program Upon Activation in Humans and Differ Energetically From Memory CD8+ T-Cells.Front Immunol. 2018 Dec 21;9:2736. doi: 10.3389/fimmu.2018.02736. eCollection 2018. Front Immunol. 2018. PMID: 30619240 Free PMC article.
-
Cardiac and Renal Comorbidities in Aging People Living With HIV.Circ Res. 2024 May 24;134(11):1636-1660. doi: 10.1161/CIRCRESAHA.124.323948. Epub 2024 May 23. Circ Res. 2024. PMID: 38781295 Free PMC article. Review.
-
Immunomonitoring via ELISPOT Assay Reveals Attenuated T-Cell Immunity to CMV in Immunocompromised Liver-Transplant Patients.Cells. 2024 Apr 24;13(9):741. doi: 10.3390/cells13090741. Cells. 2024. PMID: 38727277 Free PMC article.
-
In vitro modeling of CD8+ T cell exhaustion enables CRISPR screening to reveal a role for BHLHE40.Sci Immunol. 2023 Aug 25;8(86):eade3369. doi: 10.1126/sciimmunol.ade3369. Epub 2023 Aug 18. Sci Immunol. 2023. PMID: 37595022 Free PMC article.
References
-
- Alanio C, Lemaitre F, Law HK, Hasan M, Albert ML (2010) Enumeration of human antigen‐specific naive CD8+ T cells reveals conserved precursor frequencies. Blood 115, 3718–3725. - PubMed
-
- Appay V, Sauce D (2014) Naive T cells: the crux of cellular immune aging? Exp. Gerontol. 54, 90–93. - PubMed
-
- Appay V, Douek DC, Price DA (2008) CD8+ T cell efficacy in vaccination and disease. Nat. Med. 14, 623–628. - PubMed
-
- den Braber I, Mugwagwa T, Vrisekoop N, Westera L, Mogling R, de Boer AB, Willems N, Schrijver EH, Spierenburg G, Gaiser K, Mul E, Otto SA, Ruiter AF, Ackermans MT, Miedema F, Borghans JA, de Boer RJ, Tesselaar K (2012) Maintenance of peripheral naive T cells is sustained by thymus output in mice but not humans. Immunity 36, 288–297. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
