

Structural basis of histone H3K27 trimethylation by an active polycomb repressive complex 2
- PMID: 26472914
- PMCID: PMC5220110
- DOI: 10.1126/science.aac4383
Structural basis of histone H3K27 trimethylation by an active polycomb repressive complex 2
Abstract
Polycomb repressive complex 2 (PRC2) catalyzes histone H3K27 trimethylation (H3K27me3), a hallmark of gene silencing. Here we report the crystal structures of an active PRC2 complex of 170 kilodaltons from the yeast Chaetomium thermophilum in both basal and stimulated states, which contain Ezh2, Eed, and the VEFS domain of Suz12 and are bound to a cancer-associated inhibiting H3K27M peptide and a S-adenosyl-l-homocysteine cofactor. The stimulated complex also contains an additional stimulating H3K27me3 peptide. Eed is engulfed by a belt-like structure of Ezh2, and Suz12(VEFS) contacts both of these two subunits to confer an unusual split active SET domain for catalysis. Comparison of PRC2 in the basal and stimulated states reveals a mobile Ezh2 motif that responds to stimulation to allosterically regulate the active site.
Copyright © 2015, American Association for the Advancement of Science.
Figures

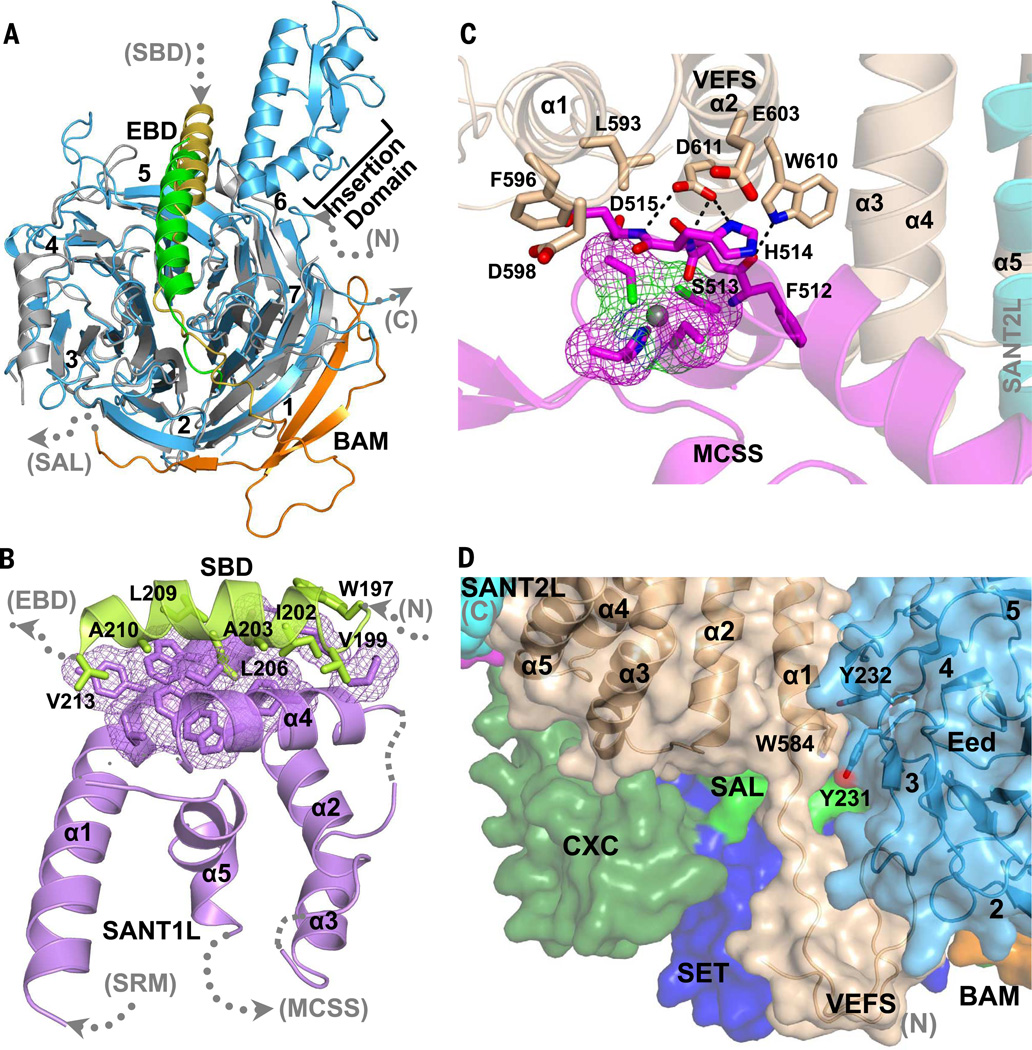
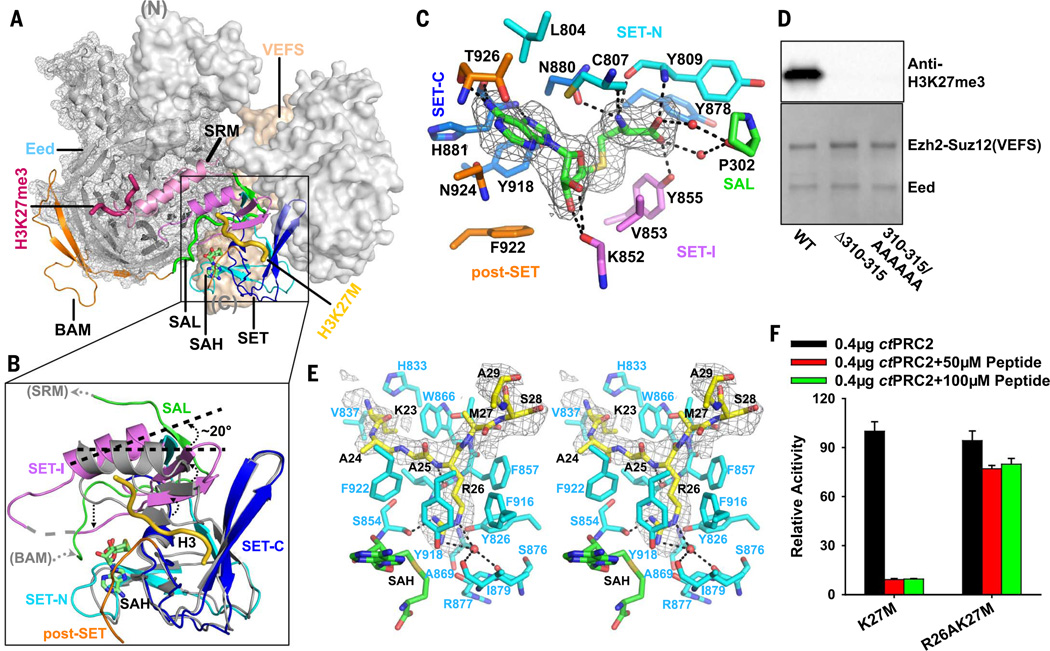
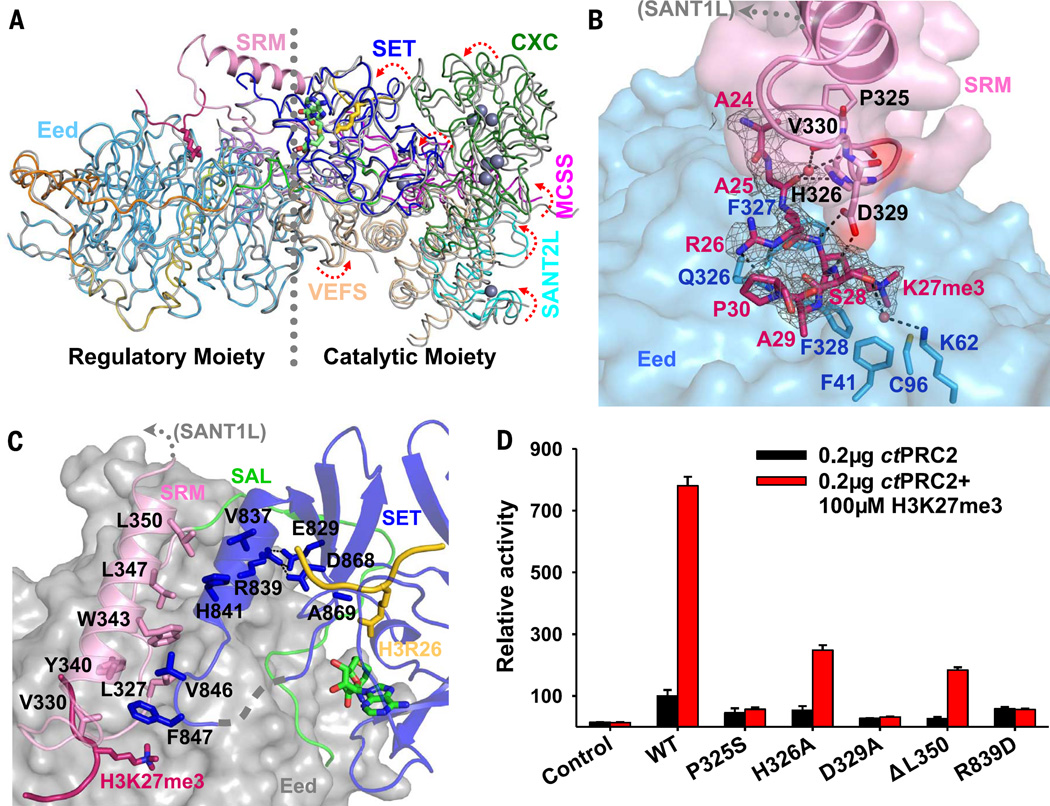
Comment in
-
STRUCTURAL BIOLOGY. Chromatin complex, crystal clear.Science. 2015 Oct 16;350(6258):278-9. doi: 10.1126/science.aad5203. Science. 2015. PMID: 26472895 No abstract available.
-
Comment on "Structural basis of histone H3K27 trimethylation by an active polycomb repressive complex 2".Science. 2016 Dec 23;354(6319):1543. doi: 10.1126/science.aaf6236. Science. 2016. PMID: 28008037
References
-
- Cao R, Zhang Y. The functions of E(Z)/EZH2-mediated methylation of lysine 27 in histone H3. Curr. Opin. Genet. Dev. 2004;14:155–164. pmid: 15196462. - PubMed
-
- Simon JA, Kingston RE. Mechanisms of polycomb gene silencing: Knowns and unknowns. Nat. Rev. Mol. Cell Biol. 2009;10:697–708. pmid: 19738629. - PubMed
-
- Delest A, Sexton T, Cavalli G. Polycomb: A paradigm for genome organization from one to three dimensions. Curr. Opin. Cell Biol. 2012;24:405–414. pmid: 22336329. - PubMed
-
- Di Croce L, Helin K. Transcriptional regulation by Polycomb group proteins. Nat. Struct. Mol. Biol. 2013;20:1147–1155. pmid: 24096405. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
