

An internal promoter underlies the difference in disease severity between N- and C-terminal truncation mutations of Titin in zebrafish
- PMID: 26473617
- PMCID: PMC4720518
- DOI: 10.7554/eLife.09406
An internal promoter underlies the difference in disease severity between N- and C-terminal truncation mutations of Titin in zebrafish
Abstract
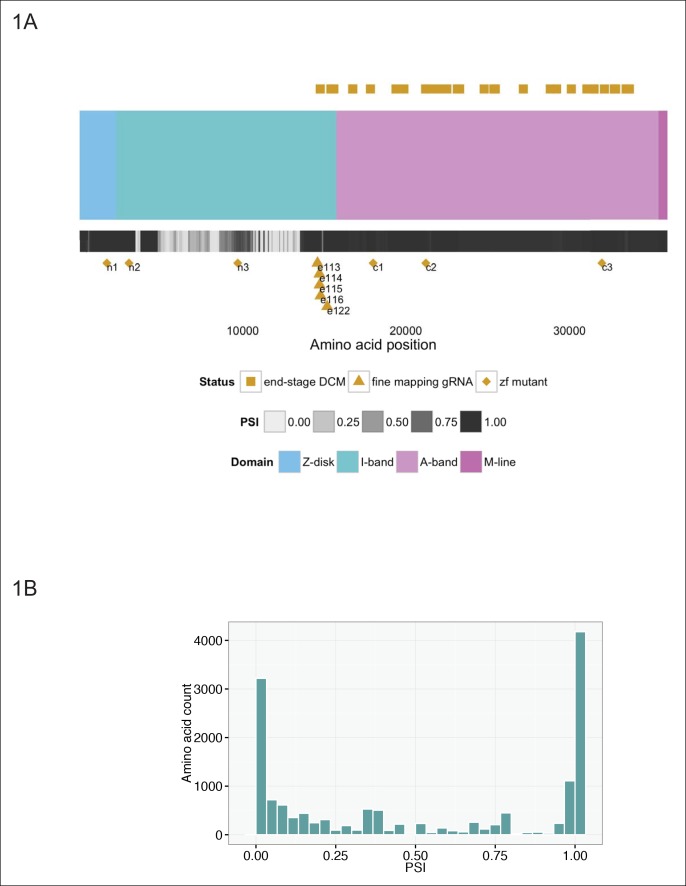
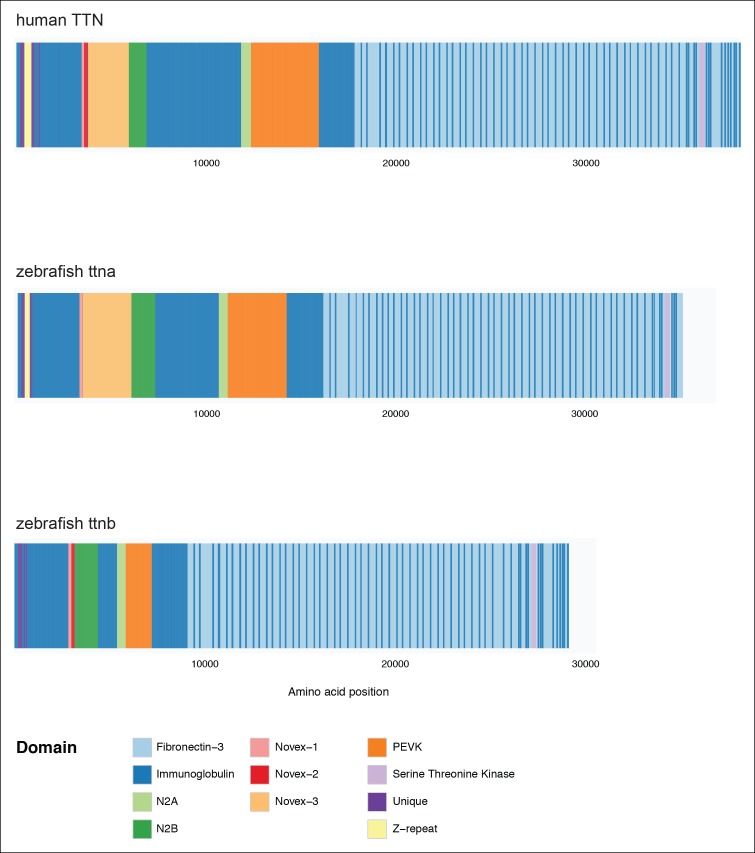
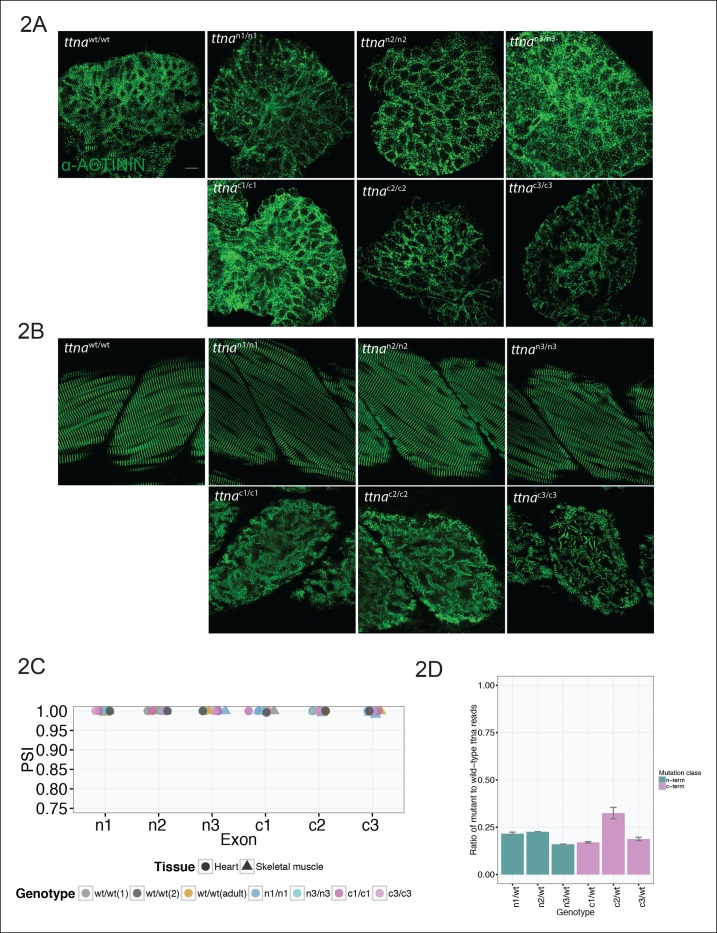
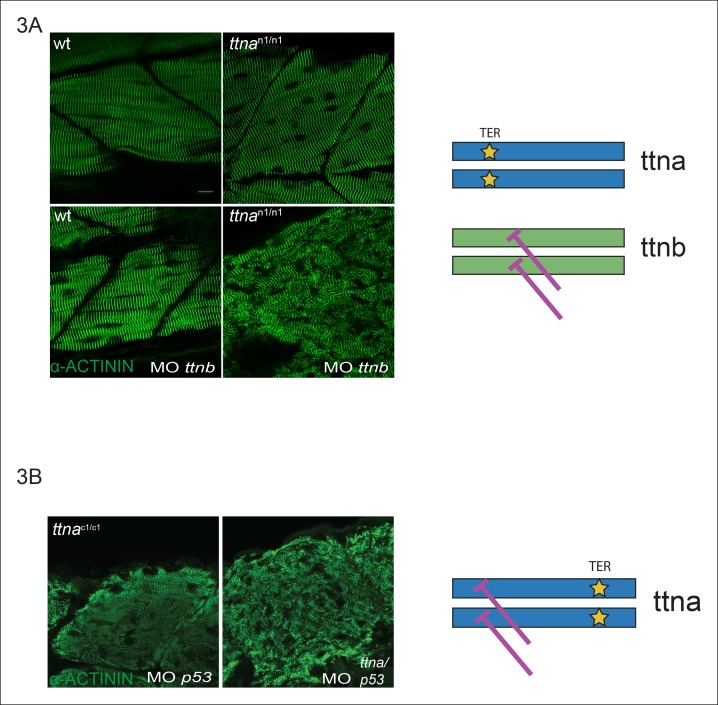
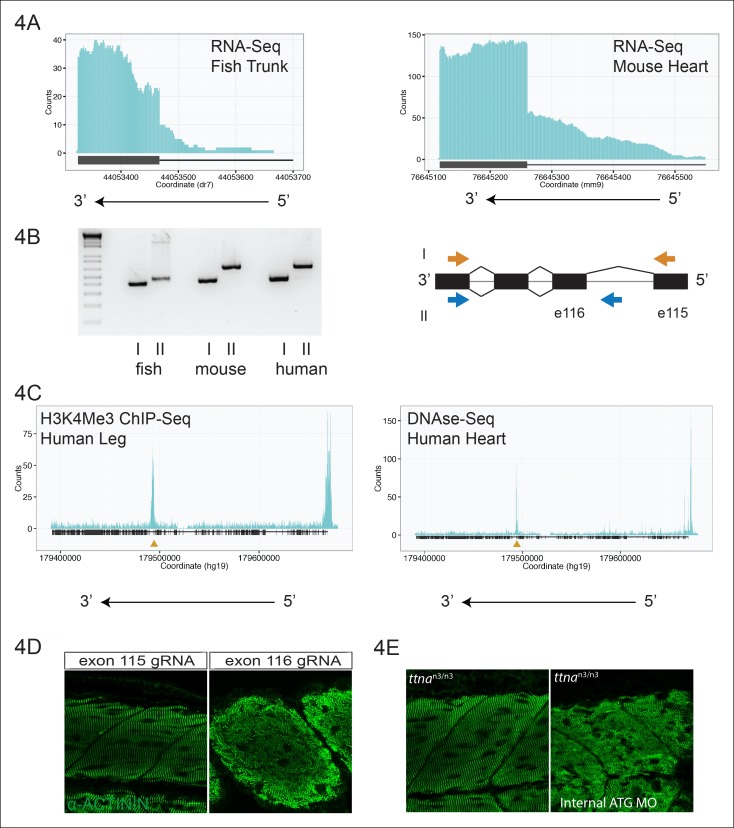
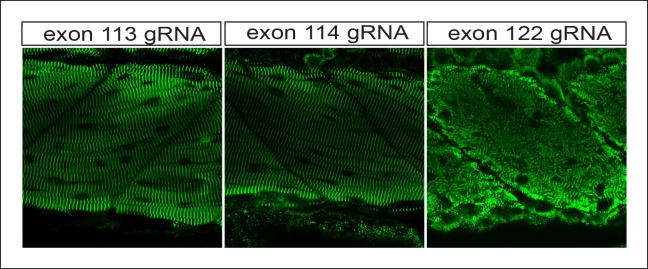
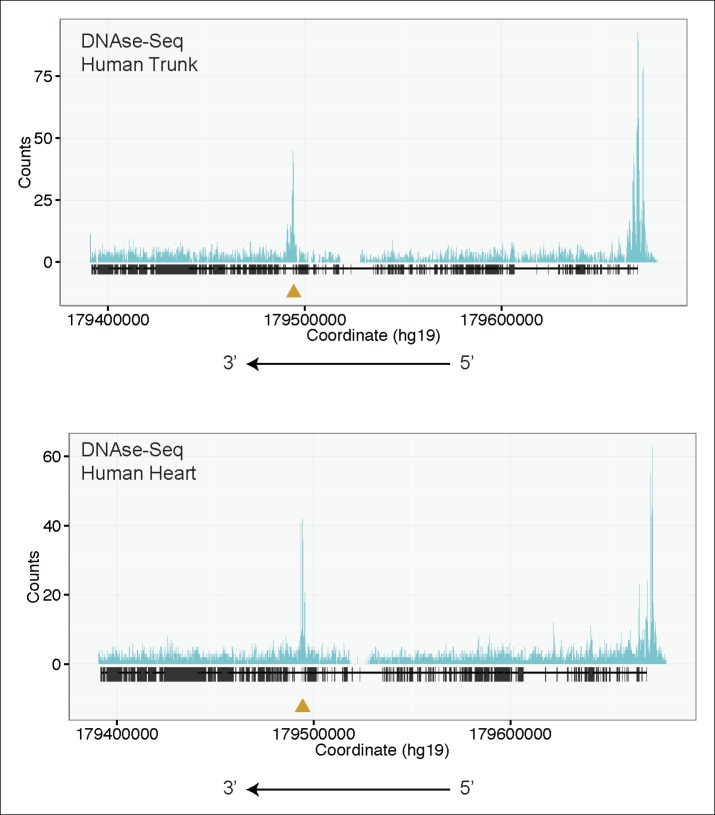
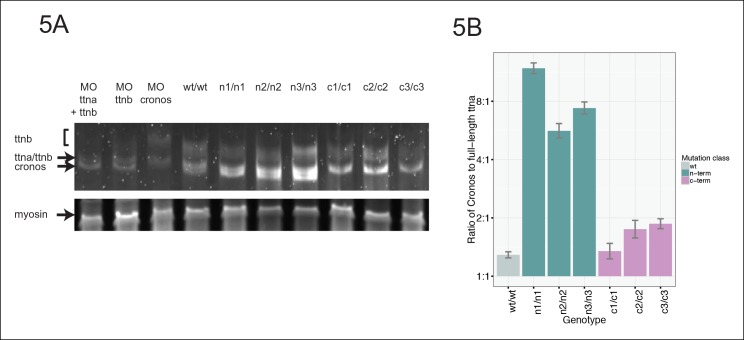
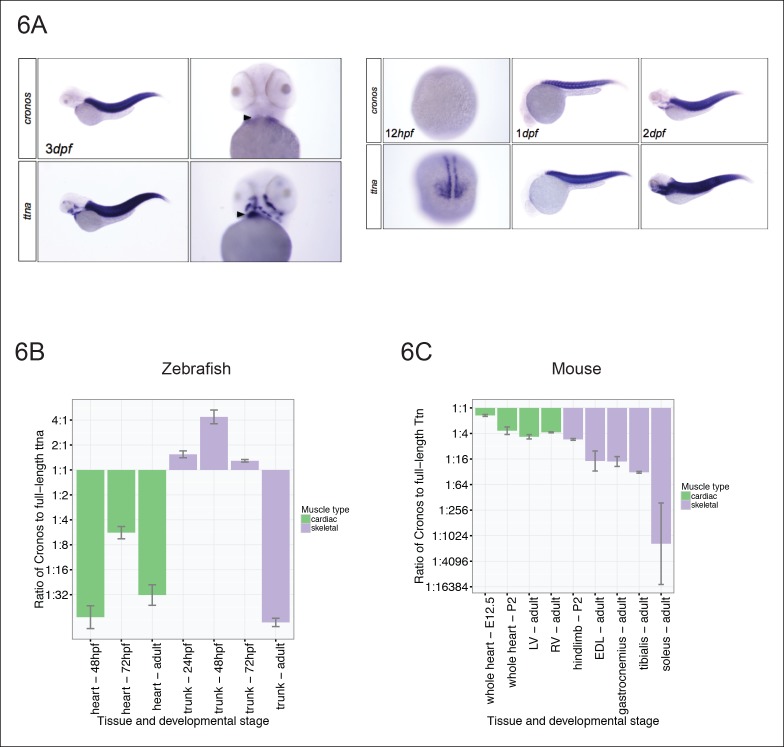
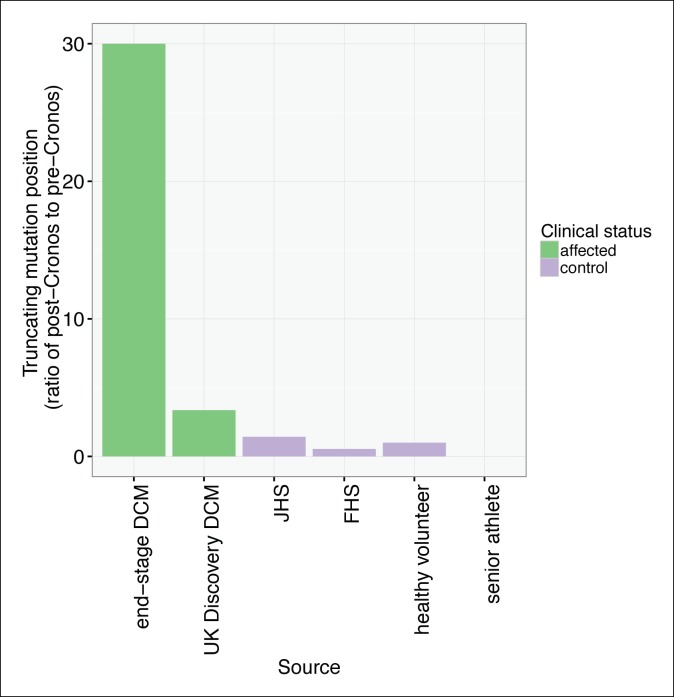
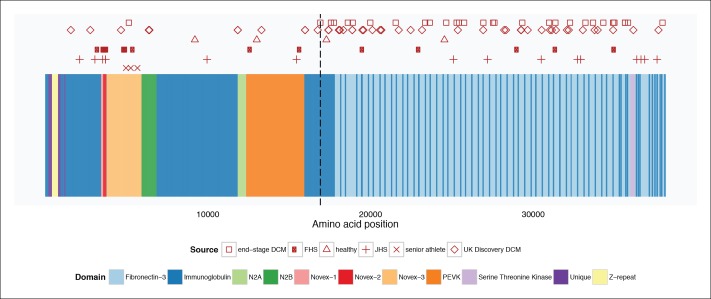
Truncating mutations in the giant sarcomeric protein Titin result in dilated cardiomyopathy and skeletal myopathy. The most severely affected dilated cardiomyopathy patients harbor Titin truncations in the C-terminal two-thirds of the protein, suggesting that mutation position might influence disease mechanism. Using CRISPR/Cas9 technology, we generated six zebrafish lines with Titin truncations in the N-terminal and C-terminal regions. Although all exons were constitutive, C-terminal mutations caused severe myopathy whereas N-terminal mutations demonstrated mild phenotypes. Surprisingly, neither mutation type acted as a dominant negative. Instead, we found a conserved internal promoter at the precise position where divergence in disease severity occurs, with the resulting protein product partially rescuing N-terminal truncations. In addition to its clinical implications, our work may shed light on a long-standing mystery regarding the architecture of the sarcomere.
Keywords: cardiomyopathy; genetics; human; human biology; medicine; mouse; sarcomere; zebrafish.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures
Similar articles
-
Alternative Splicing, Internal Promoter, Nonsense-Mediated Decay, or All Three: Explaining the Distribution of Truncation Variants in Titin.Circ Cardiovasc Genet. 2016 Oct;9(5):419-425. doi: 10.1161/CIRCGENETICS.116.001513. Epub 2016 Sep 13. Circ Cardiovasc Genet. 2016. PMID: 27625338 Free PMC article.
-
A-Band Titin Truncation in Zebrafish Causes Dilated Cardiomyopathy and Hemodynamic Stress Intolerance.Circ Genom Precis Med. 2018 Aug;11(8):e002135. doi: 10.1161/CIRCGEN.118.002135. Circ Genom Precis Med. 2018. PMID: 30354343
-
Functional analysis of titin/connectin N2-B mutations found in cardiomyopathy.J Muscle Res Cell Motil. 2005;26(6-8):367-74. doi: 10.1007/s10974-005-9018-5. J Muscle Res Cell Motil. 2005. PMID: 16465475
-
The giant titin: how to evaluate its role in cardiomyopathies.J Muscle Res Cell Motil. 2019 Jun;40(2):159-167. doi: 10.1007/s10974-019-09518-w. Epub 2019 May 30. J Muscle Res Cell Motil. 2019. PMID: 31147888 Free PMC article. Review.
-
Titin in muscular dystrophy and cardiomyopathy: Urinary titin as a novel marker.Clin Chim Acta. 2019 Aug;495:123-128. doi: 10.1016/j.cca.2019.04.005. Epub 2019 Apr 5. Clin Chim Acta. 2019. PMID: 30959043 Review.
Cited by
-
Use of animal models to understand titin physiology and pathology.J Cell Mol Med. 2022 Oct;26(20):5103-5112. doi: 10.1111/jcmm.17533. Epub 2022 Sep 6. J Cell Mol Med. 2022. PMID: 36065969 Free PMC article. Review.
-
Exon- and contraction-dependent functions of titin in sarcomere assembly.Development. 2016 Dec 15;143(24):4713-4722. doi: 10.1242/dev.139246. Epub 2016 Nov 11. Development. 2016. PMID: 27836965 Free PMC article.
-
Modeling Human Cardiac Arrhythmias: Insights from Zebrafish.J Cardiovasc Dev Dis. 2022 Jan 5;9(1):13. doi: 10.3390/jcdd9010013. J Cardiovasc Dev Dis. 2022. PMID: 35050223 Free PMC article. Review.
-
Mechanisms of TTNtv-Related Dilated Cardiomyopathy: Insights from Zebrafish Models.J Cardiovasc Dev Dis. 2021 Jan 25;8(2):10. doi: 10.3390/jcdd8020010. J Cardiovasc Dev Dis. 2021. PMID: 33504111 Free PMC article. Review.
-
Alternative Splicing, Internal Promoter, Nonsense-Mediated Decay, or All Three: Explaining the Distribution of Truncation Variants in Titin.Circ Cardiovasc Genet. 2016 Oct;9(5):419-425. doi: 10.1161/CIRCGENETICS.116.001513. Epub 2016 Sep 13. Circ Cardiovasc Genet. 2016. PMID: 27625338 Free PMC article.
References
-
- Bang M-L, Centner T, Fornoff F, Geach AJ, Gotthardt M, Mcnabb M, Witt CC, Labeit D, Gregorio CC, Granzier H, Labeit S. The complete gene sequence of titin, expression of an unusual 700-kDa titin isoform, and its interaction with obscurin identify a novel z-line to i-band linking system. Circulation Research. 2001;89:1065–1072. doi: 10.1161/hh2301.100981. - DOI - PubMed
-
- Carmignac V, Salih MA, Quijano-Roy S, Marchand S, Al Rayess MM, Mukhtar MM, Urtizberea JA, Labeit S, Guicheney P, Leturcq F, Gautel M, Fardeau M, Campbell KP, Richard I, Estournet B, Ferreiro A. C-terminal titin deletions cause a novel early-onset myopathy with fatal cardiomyopathy. Annals of Neurology. 2007;61:340–351. doi: 10.1002/ana.21089. - DOI - PubMed
-
- Ceyhan-Birsoy O, Agrawal PB, Hidalgo C, Schmitz-Abe K, DeChene ET, Swanson LC, Soemedi R, Vasli N, Iannaccone ST, Shieh PB, Shur N, Dennison JM, Lawlor MW, Laporte J, Markianos K, Fairbrother WG, Granzier H, Beggs AH. Recessive truncating titin gene, TTN, mutations presenting as centronuclear myopathy. Neurology. 2013;81:1205–1214. doi: 10.1212/WNL.0b013e3182a6ca62. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
