

Lethal Neonatal Progression of Fetal Cardiomegaly Associated to ACAD9 Deficiency
- PMID: 26475292
- PMCID: PMC5059192
- DOI: 10.1007/8904_2015_499
Lethal Neonatal Progression of Fetal Cardiomegaly Associated to ACAD9 Deficiency
Abstract
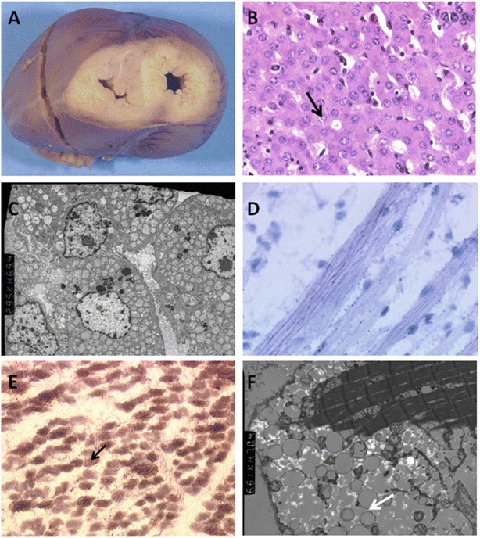
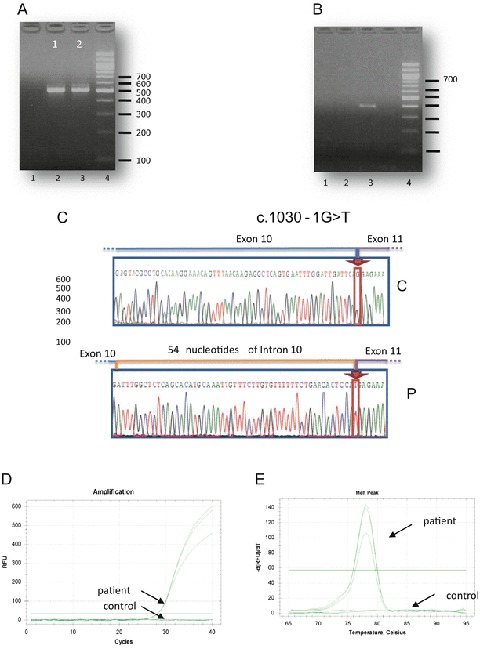
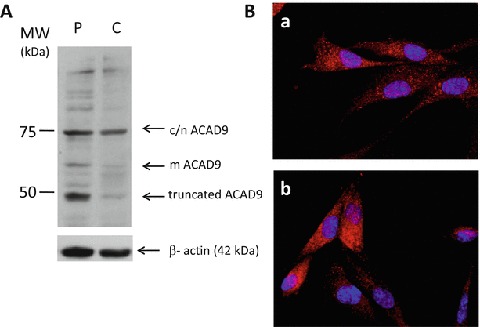
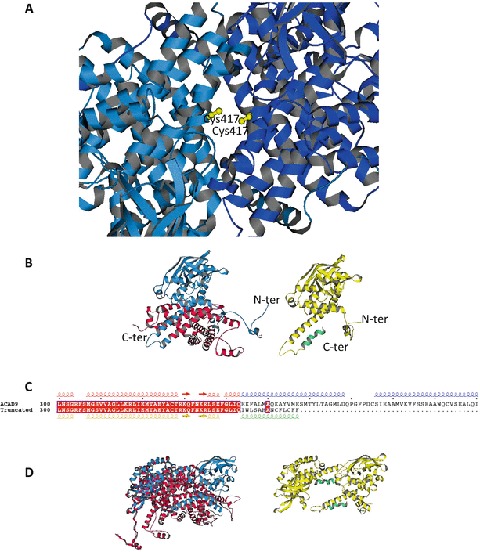
ACAD9 (acyl-CoA dehydrogenase 9) is an essential factor for the mitochondrial respiratory chain complex I assembly. ACAD9, a member of acyl-CoA dehydrogenase family, has high homology with VLCAD (very long-chain acyl-CoA dehydrogenase) and harbors a homodimer structure. Recently, patients with ACAD9 deficiency have been described with a wide clinical spectrum ranging from severe lethal form to moderate form with exercise intolerance.We report here a prenatal presentation with intrauterine growth retardation and cardiomegaly, with a fatal outcome shortly after birth. Compound heterozygous mutations, a splice-site mutation - c.1030-1G>T and a missense mutation - c.1249C>T; p.Arg417Cys, were identified in the ACAD9 gene. Their effect on protein structure and expression level was investigated. Protein modeling suggested a functional effect of the c.1030-1G>T mutation generating a non-degraded truncated protein and the p.Arg417Cys, creating an aberrant dimer. Our results underscore the crucial role of ACAD9 protein for cardiac function.
Keywords: ACAD9; Fetal cardiomegaly; Mitochondrial respiratory chain; β-Oxidation.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
