

On intrinsic stress fiber contractile forces in semilunar heart valve interstitial cells using a continuum mixture model
- PMID: 26476967
- PMCID: PMC4698364
- DOI: 10.1016/j.jmbbm.2015.09.027
On intrinsic stress fiber contractile forces in semilunar heart valve interstitial cells using a continuum mixture model
Abstract
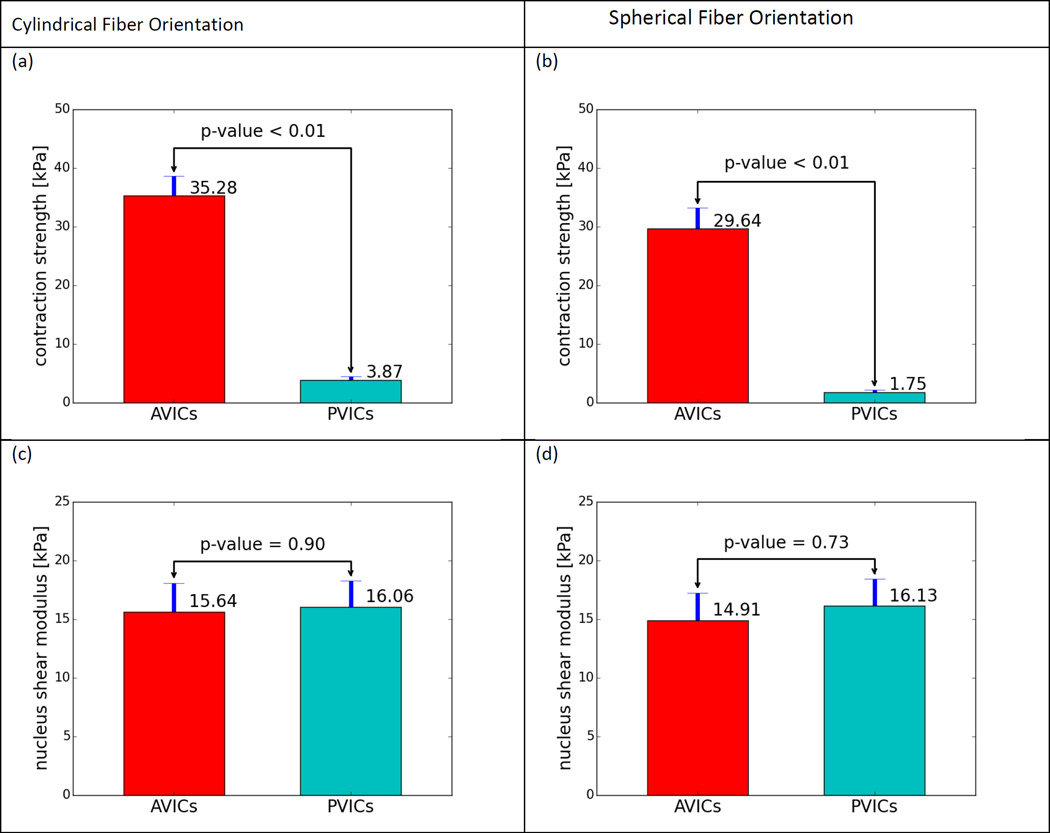
Heart valve interstitial cells (VICs) play a critical role in the maintenance and pathophysiology of heart valve tissues. Normally quiescent in the adult, VICs can become activated in periods of growth and disease. When activated, VICs exhibit increased levels of cytokines and extracellular matrix (ECM) synthesis, and upregulated expression and strong contraction of α-smooth muscle actin (α-SMA) fibers. However, it remains unknown how expression and contraction of the α-SMA fibers, which vary among different VIC types, contribute to the overall VIC mechanical responses, including the nucleus and cytoskeleton contributions. In the present study, we developed a novel solid-mixture model for VIC biomechanical behavior that incorporated 1) the underlying cytoskeletal network, 2) the oriented α-SMA stress fibers with passive elastic and active contractile responses, 3) a finite deformable elastic nucleus. We implemented the model in a full 3D finite element simulation of a VIC based on known geometry. Moreover, we examined the respective mechanical responses of aortic and pulmonary VICs (AVICs and PVICs, respectively), which are known to have different levels of α-SMA expression levels and contractile behaviors. To calibrate the model, we simulated the combined mechanical responses of VICs in both micropipette aspiration (MA) and atomic force microscopy (AFM) experiments. These two states were chosen as the VICs were under significantly different mechanical loading conditions and activation states, with the α-SMA fibers inactivated in the MA studies while fully activated in the AFM studies. We also used the AFM to study the mechanical property of the nucleus. Our model predicted that the substantial differences found in stiffening of the AVIC compared to the PVICs was due to a 9 to 16 times stronger intrinsic AVIC α-SMA stress fiber contractile force. Model validation was done by simulating a traction force microscopy experiment to estimate the forces the VICs exert on the underlying substrate, and found good agreement with reported traction force microscopy results. Further, estimated nuclear stiffness for both AVICs and PVICs were similar and comparable to the literature, and were both unaffected by VIC activation level. These results suggest substantial functional differences between AVICs and PVICs at the subcellular level. Moreover, this first VIC computational biomechanical model is but a first step in developing a comprehensive, integrated view of the VIC pathophysiology and interactions with the valve ECM micro-environment based on simulation technologies.
Keywords: Active contraction; Alpha-SMA stress fibers; Atomic force microscopy; Finite element method; Heart valves; Micropipette aspiration; Valve interstitial cell.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures











References
-
- Sacks MS, Schoen FJ, Mayer JE. Bioengineering challenges for heart valve tissue engineering. Annual Review of Biomedical Engineering. 2009;11:289–313. - PubMed
-
- Schoen FJ. Evolving concepts of cardiac valve dynamics: the continuum of development, functional structure, pathobiology, and tissue engineering. Circulation. 2008;118(18):1864–1880. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
