

Functional Integration of Adult-Born Hippocampal Neurons after Traumatic Brain Injury(1,2,3)
- PMID: 26478908
- PMCID: PMC4603252
- DOI: 10.1523/ENEURO.0056-15.2015
Functional Integration of Adult-Born Hippocampal Neurons after Traumatic Brain Injury(1,2,3)
Abstract
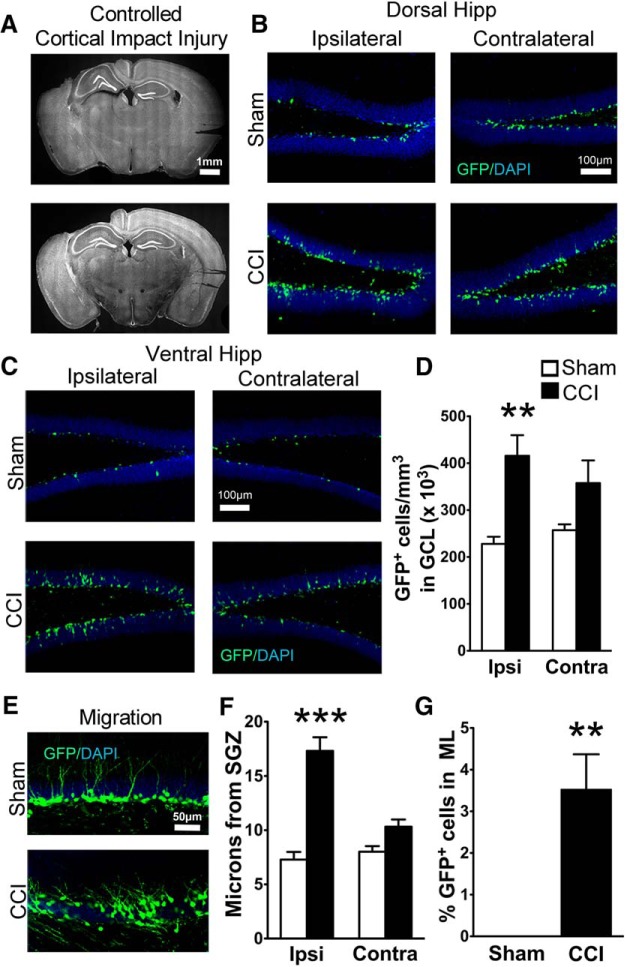
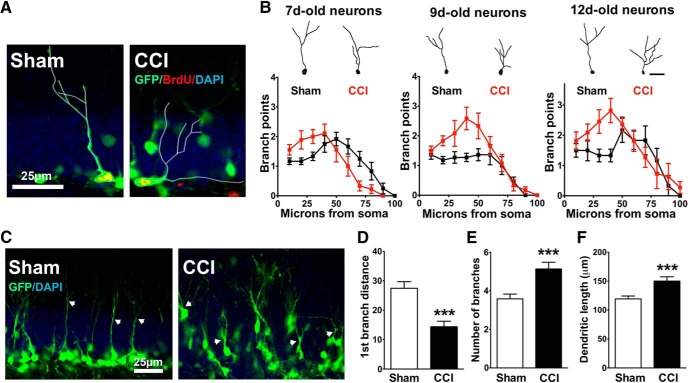
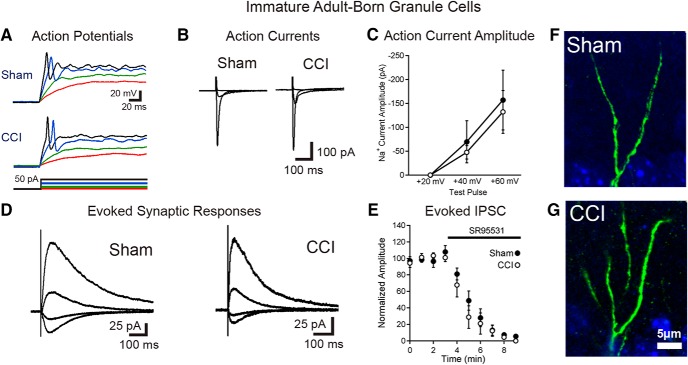
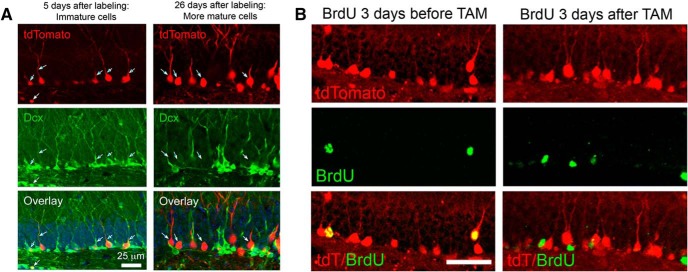
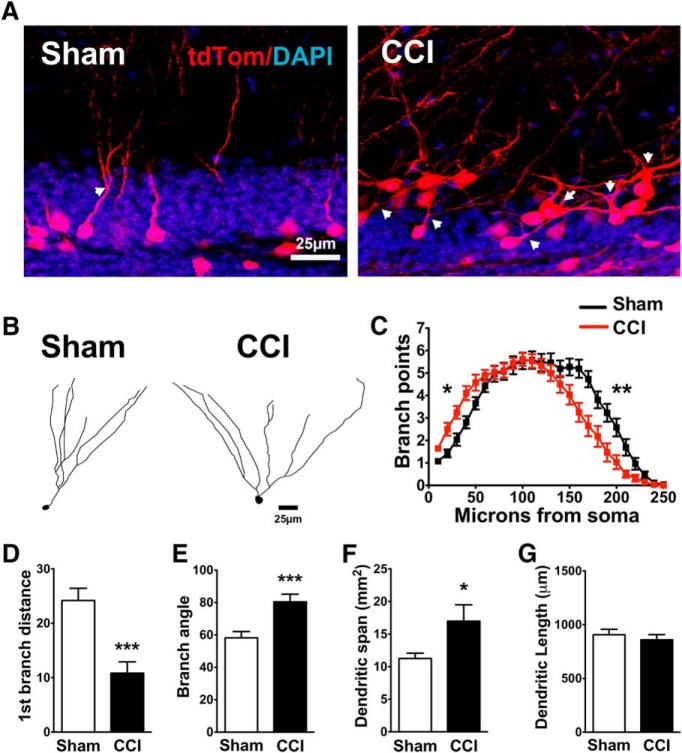
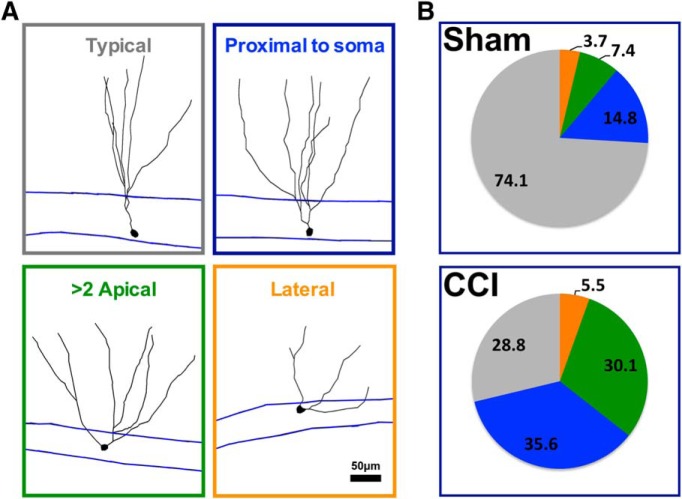
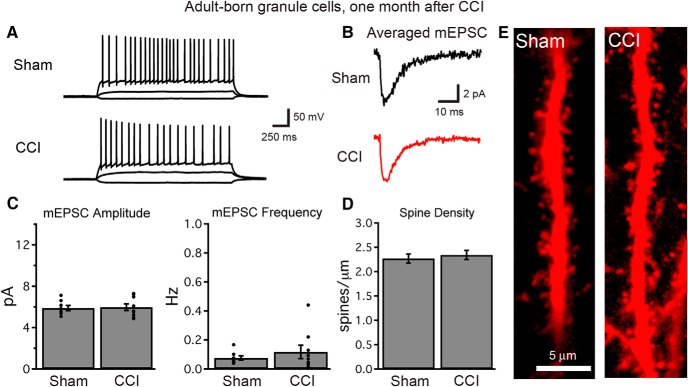
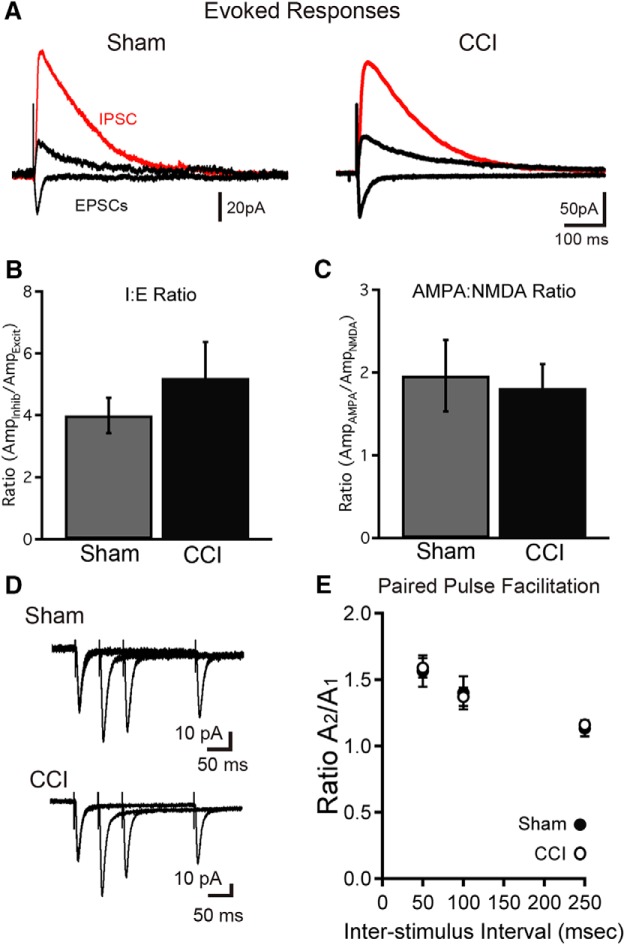
Traumatic brain injury (TBI) increases hippocampal neurogenesis, which may contribute to cognitive recovery after injury. However, it is unknown whether TBI-induced adult-born neurons mature normally and functionally integrate into the hippocampal network. We assessed the generation, morphology, and synaptic integration of new hippocampal neurons after a controlled cortical impact (CCI) injury model of TBI. To label TBI-induced newborn neurons, we used 2-month-old POMC-EGFP mice, which transiently and specifically express EGFP in immature hippocampal neurons, and doublecortin-CreER(T2) transgenic mice crossed with Rosa26-CAG-tdTomato reporter mice, to permanently pulse-label a cohort of adult-born hippocampal neurons. TBI increased the generation, outward migration, and dendritic complexity of neurons born during post-traumatic neurogenesis. Cells born after TBI had profound alterations in their dendritic structure, with increased dendritic branching proximal to the soma and widely splayed dendritic branches. These changes were apparent during early dendritic outgrowth and persisted as these cells matured. Whole-cell recordings from neurons generated during post-traumatic neurogenesis demonstrate that they are excitable and functionally integrate into the hippocampal circuit. However, despite their dramatic morphologic abnormalities, we found no differences in the rate of their electrophysiological maturation, or their overall degree of synaptic integration when compared to age-matched adult-born cells from sham mice. Our results suggest that cells born after TBI participate in information processing, and receive an apparently normal balance of excitatory and inhibitory inputs. However, TBI-induced changes in their anatomic localization and dendritic projection patterns could result in maladaptive network properties.
Keywords: adult neurogenesis; functional integration; hippocampus; maturation; synaptic integration; traumatic brain injury.
Conflict of interest statement
The authors declare no competing financial interests.
Figures

References
-
- Cameron HA, Woolley CS, McEwen BS, Gould E (1993) Differentiation of newly born neurons and glia in the dentate gyrus of the adult rat. Neuroscience 56:337-344. - PubMed
-
- Carlson SW, Madathil SK, Sama DM, Gao X, Chen J, Saatman KE (2014) Conditional overexpression of insulin-like growth factor-1 enhances hippocampal neurogenesis and restores immature neuron dendritic processes after traumatic brain injury. J Neuropathol Exp Neurol 73:734-746. 10.1097/NEN.0000000000000092 - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous