

The kinase inhibitor D11 induces caspase-mediated cell death in cancer cells resistant to chemotherapeutic treatment
- PMID: 26480820
- PMCID: PMC4612421
- DOI: 10.1186/s13046-015-0234-6
The kinase inhibitor D11 induces caspase-mediated cell death in cancer cells resistant to chemotherapeutic treatment
Abstract
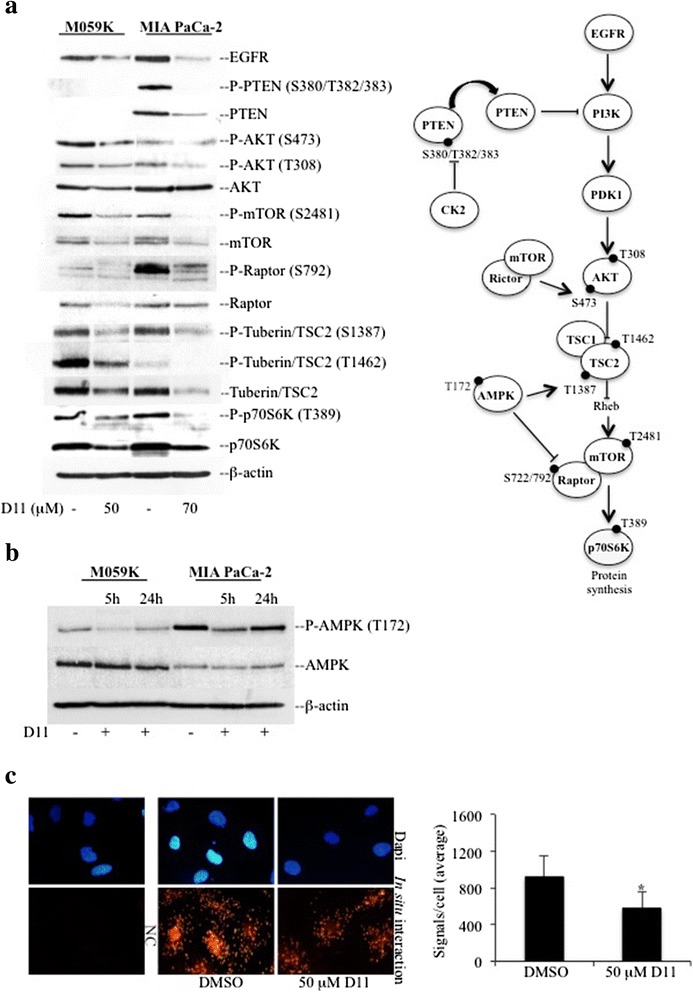
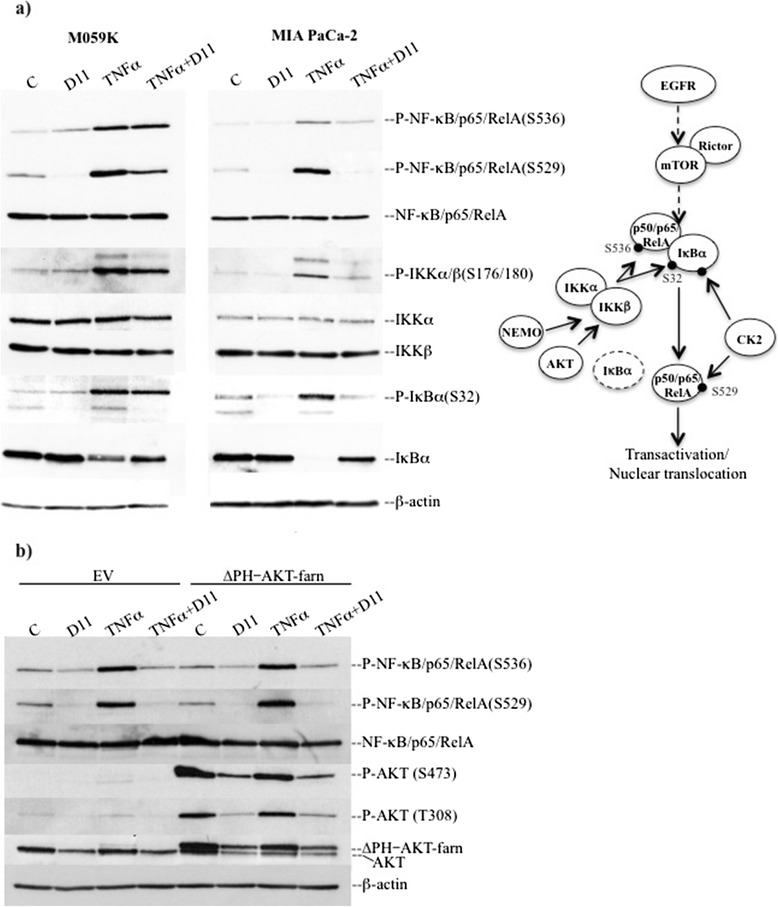
Background: Multi-drug resistance and predisposition to metastasize are major clinical problems in cancer treatment. Malignant primary brain tumor and pancreatic cancer are two well-known examples of malignant tumors resistant to conventional therapies where aberrant EGFR-mediated and NF-κB signal transduction pathways are likely to play an important role. We have recently identified 1,3-Dichloro-6-[(E)-((4-methoxyphenyl)imino)methyl] diben-zo(b,d) furan-2,7-diol (D11) as a potent and selective inhibitor of CK2 a serine/threonine protein kinase that modulates the aforementioned signaling cascades.
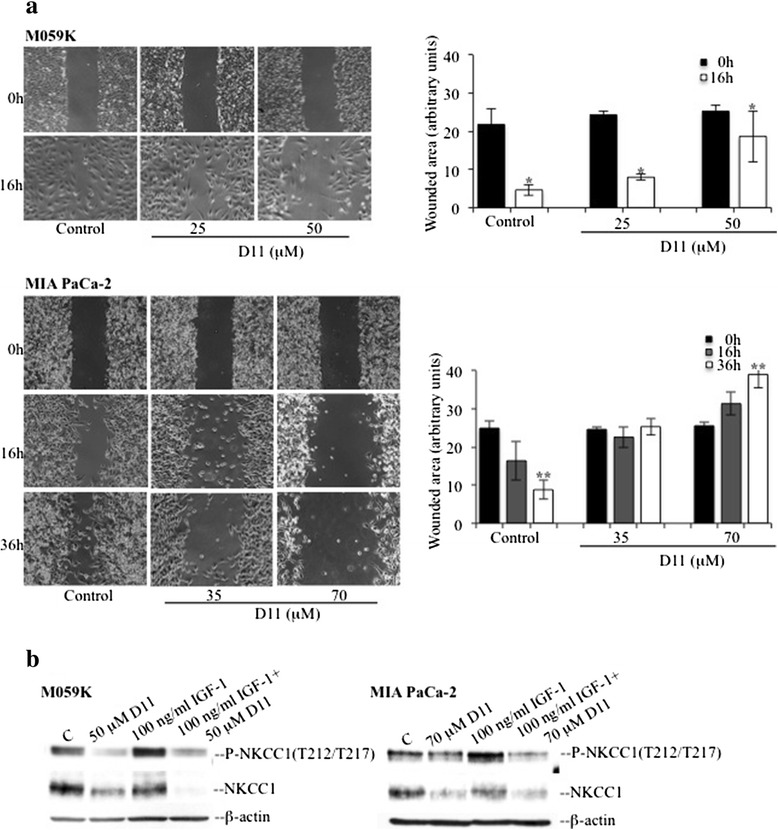
Methods: Human cancer cell lines (glioblastoma and pancreatic adenocarcinoma) resistant to conventional chemotherapeutic agents were incubated with increasing concentrations of D11 for variable amounts of time. Cell viability, cell death and effects on major signal transduction pathways deregulated in cancer cells were analyzed by ELISA, FACS and Western blot-based assays, respectively. Moreover, effects on cell migration and in cell protein-protein association were investigated by wound-healing and in situ proximity ligation assays, respectively.
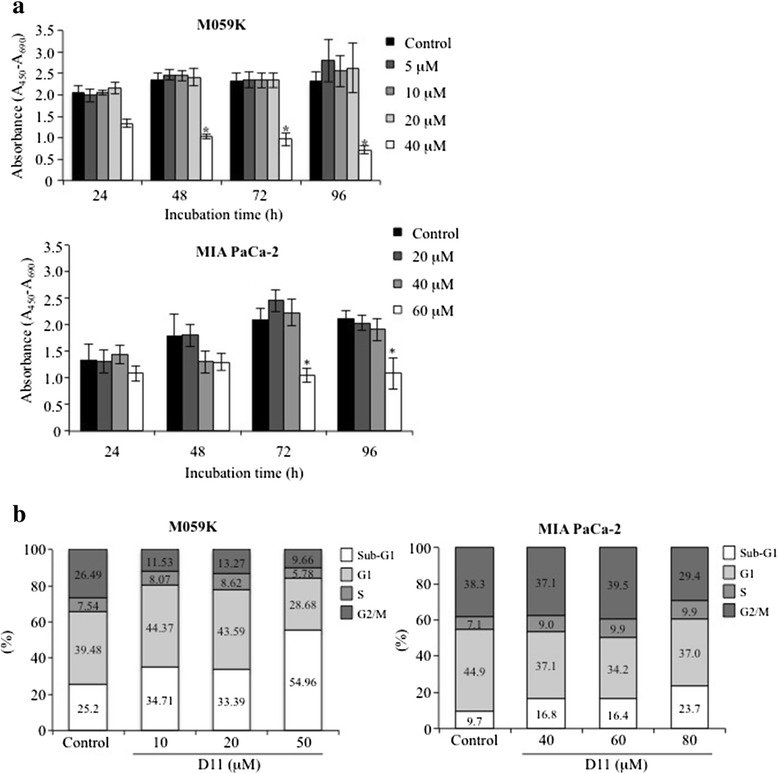
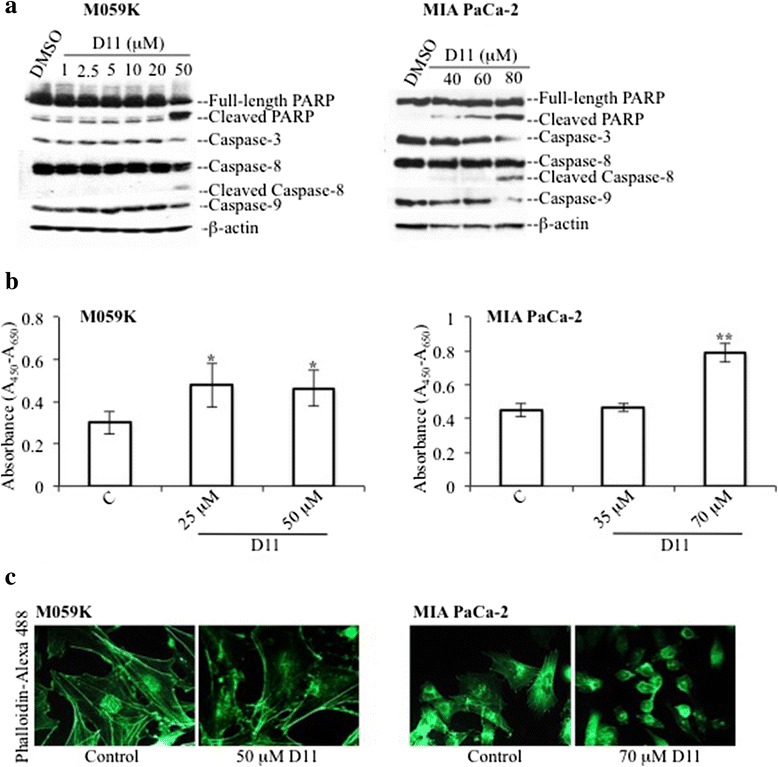
Results: We show here, that D11 treatment leads to i) significant caspase-mediated apoptotic cell death, ii) down-regulation of EGFR expression and iii) inhibition of NF-κB transcriptional activity. Furthermore, cell exposure to D11 results in impaired cell migration and correlates with reduced expression of the ion co-transporter and cell volume regulator Na(+)-K(+)-2Cl(-) (NKCC1).
Conclusions: Data reported here underline the therapeutic potential of D11 with respect to certain types of cancer that carry aberrant intracellular signaling cascades and/or exhibit sustained cell migration and suggest a new therapeutic strategy against chemotherapy resistance.
Figures

References
-
- Cancer Genome Atlas Research Network. Kandoth C, Schultz N, Cherniack AD, Akbani R, Liu Y, Shen H, Robertson AG, Pashtan I, Shen R, Benz CC, Yau C, Laird PW, Ding L, Zhang W, Mills GB, Kucherlapati R, Mardis ER, Levine DA. Integrated genomic characterization of endometrial carcinoma. Nature. 2013;497:67–73. doi: 10.1038/nature12113. - DOI - PMC - PubMed
-
- Akimoto T, Hunter NR, Buchmiller L, Mason K, Ang KK, Milas L. Inverse relationship between epidermal growth factor receptor expression and radiocurability of murine carcinomas. Clin Cancer Res. 1999;5:2884–90. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
