

Paramutation in Drosophila Requires Both Nuclear and Cytoplasmic Actors of the piRNA Pathway and Induces Cis-spreading of piRNA Production
- PMID: 26482790
- PMCID: PMC4676525
- DOI: 10.1534/genetics.115.180307
Paramutation in Drosophila Requires Both Nuclear and Cytoplasmic Actors of the piRNA Pathway and Induces Cis-spreading of piRNA Production
Abstract
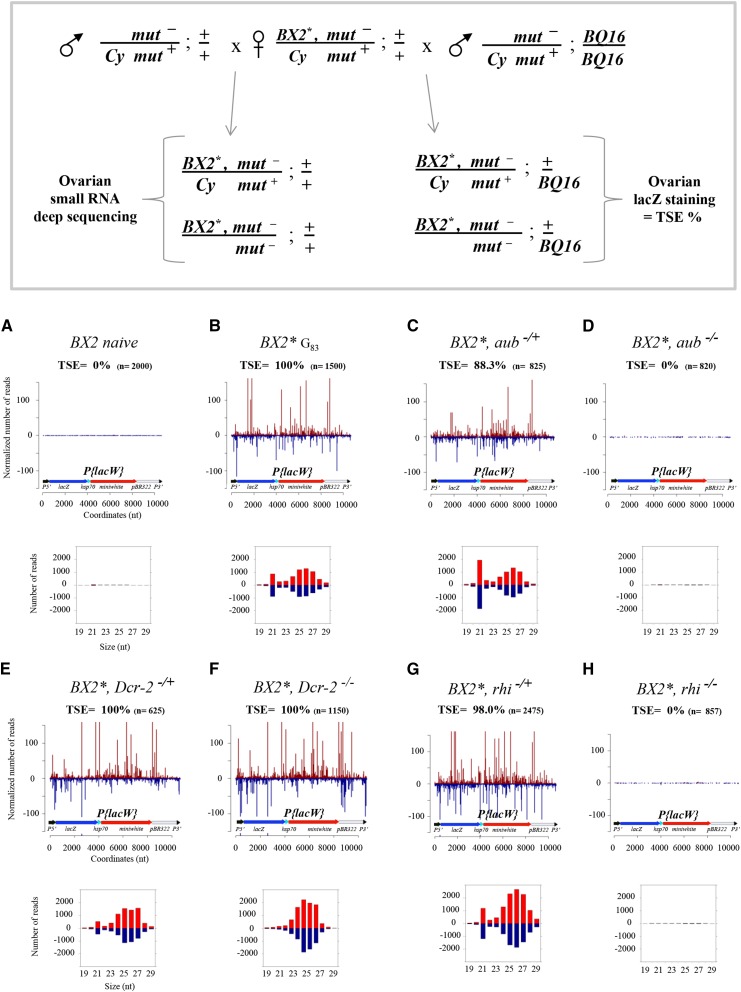
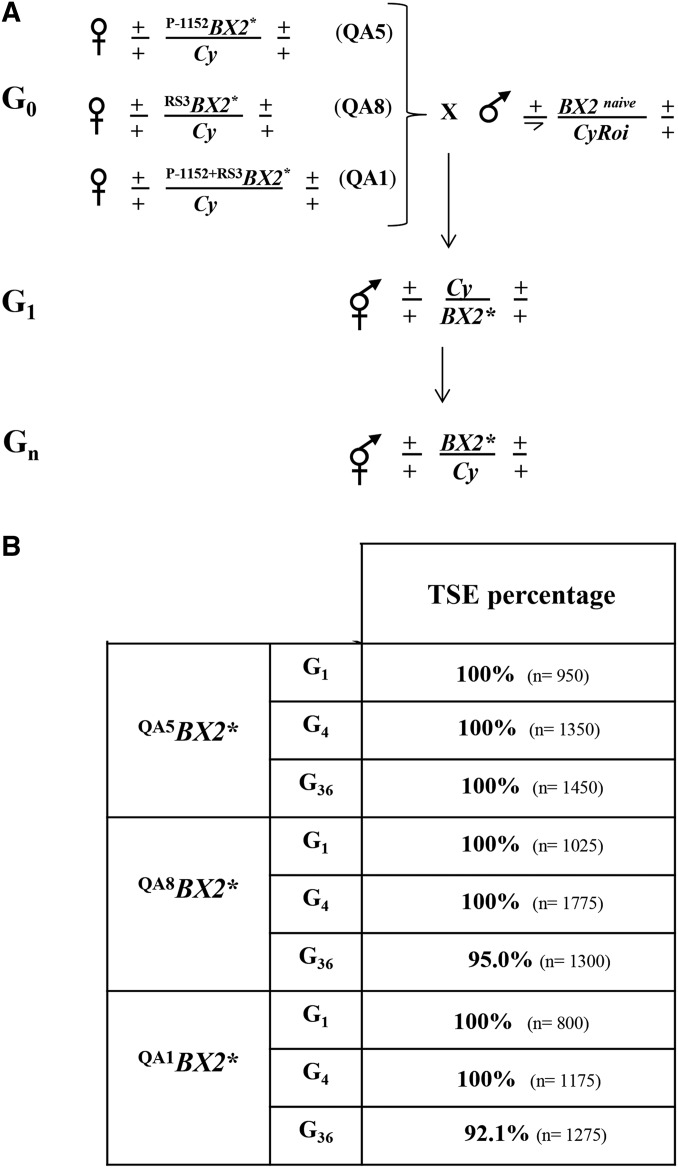
Transposable element activity is repressed in the germline in animals by PIWI-interacting RNAs (piRNAs), a class of small RNAs produced by genomic loci mostly composed of TE sequences. The mechanism of induction of piRNA production by these loci is still enigmatic. We have shown that, in Drosophila melanogaster, a cluster of tandemly repeated P-lacZ-white transgenes can be activated for piRNA production by maternal inheritance of a cytoplasm containing homologous piRNAs. This activated state is stably transmitted over generations and allows trans-silencing of a homologous transgenic target in the female germline. Such an epigenetic conversion displays the functional characteristics of a paramutation, i.e., a heritable epigenetic modification of one allele by the other. We report here that piRNA production and trans-silencing capacities of the paramutated cluster depend on the function of the rhino, cutoff, and zucchini genes involved in primary piRNA biogenesis in the germline, as well as on that of the aubergine gene implicated in the ping-pong piRNA amplification step. The 21-nt RNAs, which are produced by the paramutated cluster, in addition to 23- to 28-nt piRNAs are not necessary for paramutation to occur. Production of these 21-nt RNAs requires Dicer-2 but also all the piRNA genes tested. Moreover, cytoplasmic transmission of piRNAs homologous to only a subregion of the transgenic locus can generate a strong paramutated locus that produces piRNAs along the whole length of the transgenes. Finally, we observed that maternally inherited transgenic small RNAs can also impact transgene expression in the soma. In conclusion, paramutation involves both nuclear (Rhino, Cutoff) and cytoplasmic (Aubergine, Zucchini) actors of the piRNA pathway. In addition, since it is observed between nonfully homologous loci located on different chromosomes, paramutation may play a crucial role in epigenome shaping in Drosophila natural populations.
Keywords: Drosophila; gene regulation; mobile DNA; noncoding small RNAs; trans-generational epigenetics.
Copyright © 2015 by the Genetics Society of America.
Figures




Similar articles
-
Paramutation in Drosophila linked to emergence of a piRNA-producing locus.Nature. 2012 Oct 4;490(7418):112-5. doi: 10.1038/nature11416. Epub 2012 Aug 26. Nature. 2012. PMID: 22922650
-
Paramutation phenomena in non-vertebrate animals.Semin Cell Dev Biol. 2015 Aug;44:39-46. doi: 10.1016/j.semcdb.2015.08.009. Epub 2015 Aug 28. Semin Cell Dev Biol. 2015. PMID: 26318740 Review.
-
piRNAs and epigenetic conversion in Drosophila.Fly (Austin). 2013 Oct-Dec;7(4):237-41. doi: 10.4161/fly.26522. Epub 2013 Oct 2. Fly (Austin). 2013. PMID: 24088599 Free PMC article.
-
Profiles of piRNA abundances at emerging or established piRNA loci are determined by local DNA sequences.RNA Biol. 2013 Aug;10(8):1233-9. doi: 10.4161/rna.25756. Epub 2013 Jul 15. RNA Biol. 2013. PMID: 23880829 Free PMC article.
-
piRNA pathway and the potential processing site, the nuage, in the Drosophila germline.Dev Growth Differ. 2012 Jan;54(1):66-77. doi: 10.1111/j.1440-169x.2011.01316.x. Dev Growth Differ. 2012. PMID: 23741748 Review.
Cited by
-
Nuclear Argonaute Piwi Gene Mutation Affects rRNA by Inducing rRNA Fragment Accumulation, Antisense Expression, and Defective Processing in Drosophila Ovaries.Int J Mol Sci. 2020 Feb 7;21(3):1119. doi: 10.3390/ijms21031119. Int J Mol Sci. 2020. PMID: 32046213 Free PMC article.
-
Modeling early germline immunization after horizontal transfer of transposable elements reveals internal piRNA cluster heterogeneity.BMC Biol. 2023 May 24;21(1):117. doi: 10.1186/s12915-023-01616-z. BMC Biol. 2023. PMID: 37226160 Free PMC article.
-
piRNA Biogenesis in Drosophila melanogaster.Trends Genet. 2017 Nov;33(11):882-894. doi: 10.1016/j.tig.2017.09.002. Epub 2017 Sep 27. Trends Genet. 2017. PMID: 28964526 Free PMC article. Review.
-
Maternally inherited piRNAs direct transient heterochromatin formation at active transposons during early Drosophila embryogenesis.Elife. 2021 Jul 8;10:e68573. doi: 10.7554/eLife.68573. Elife. 2021. PMID: 34236313 Free PMC article.
-
Spoink, a LTR retrotransposon, invaded D. melanogaster populations in the 1990s.PLoS Genet. 2024 Mar 26;20(3):e1011201. doi: 10.1371/journal.pgen.1011201. eCollection 2024 Mar. PLoS Genet. 2024. PMID: 38530818 Free PMC article.
References
-
- Antoniewski C., 2014. Computing siRNA and piRNA overlap signatures. Methods Mol. Biol. 1173: 135–146. - PubMed
-
- Biemont C., Ronsseray S., Anxolabehere D., Izaabel H., Gautier C., 1990. Localization of P elements, copy number regulation, and cytotype determination in Drosophila melanogaster. Genet. Res. 56: 3–14. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
