

CaV3.2 T-type channels mediate Ca²⁺ entry during oocyte maturation and following fertilization
- PMID: 26483387
- PMCID: PMC4712821
- DOI: 10.1242/jcs.180026
CaV3.2 T-type channels mediate Ca²⁺ entry during oocyte maturation and following fertilization
Abstract
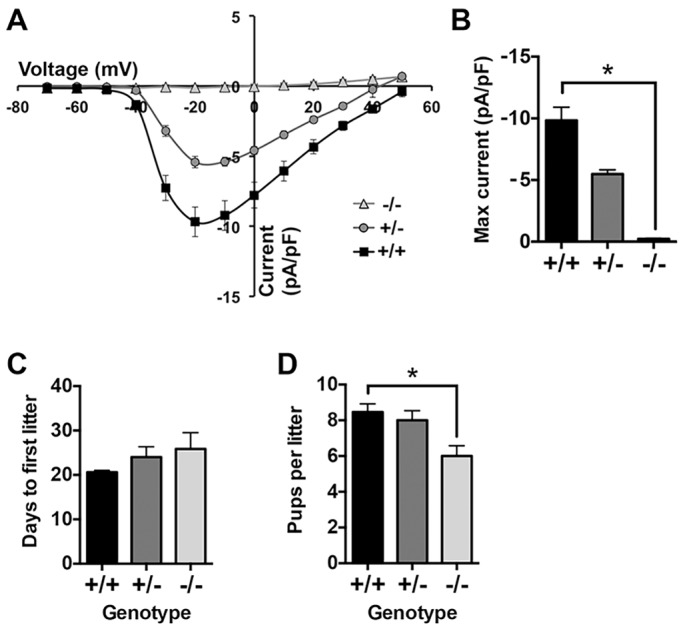
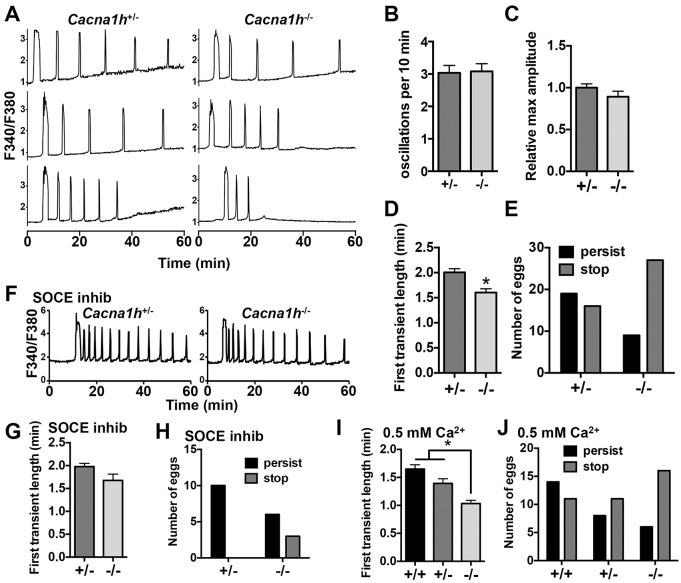
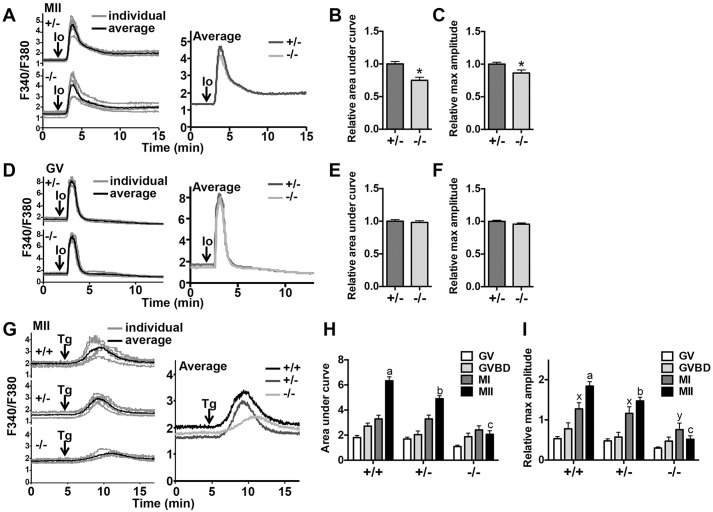
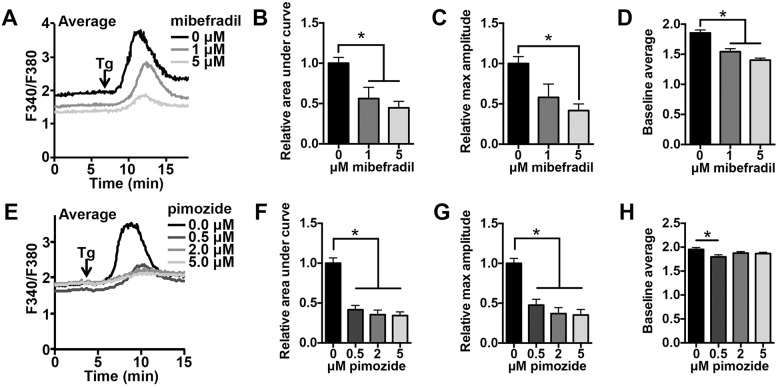
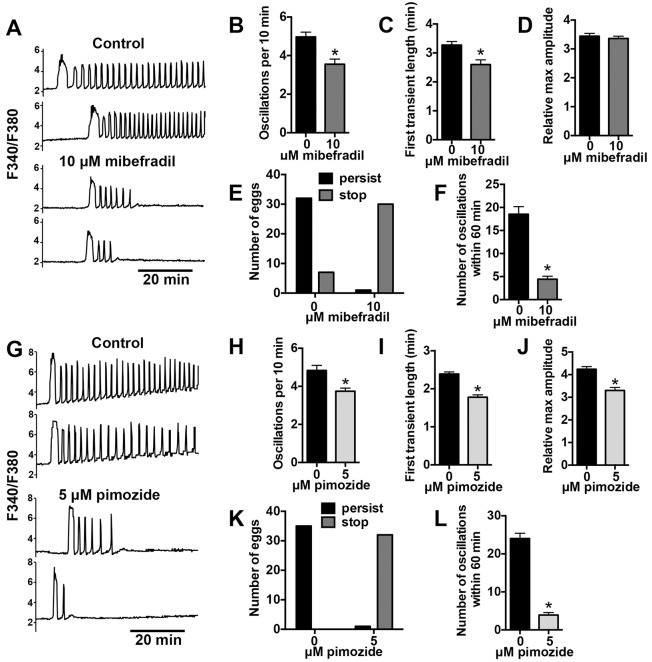
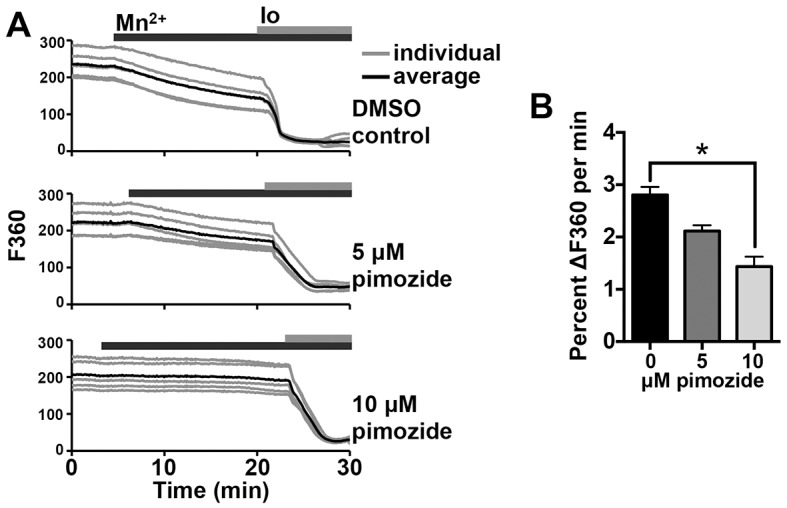
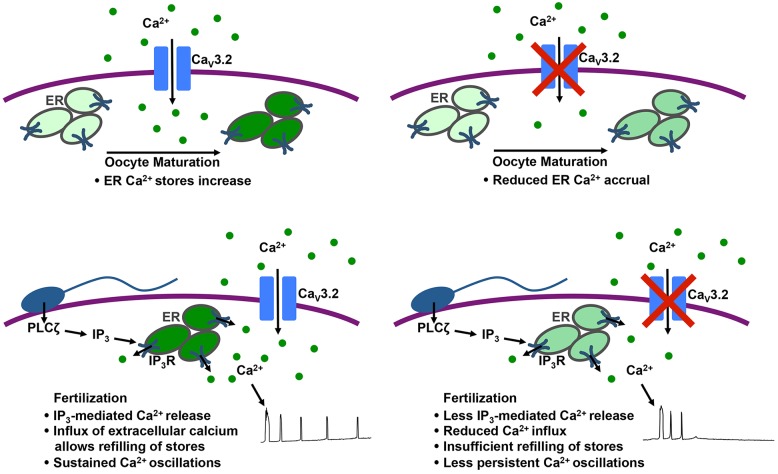
Initiation of mouse embryonic development depends upon a series of fertilization-induced rises in intracellular Ca(2+). Complete egg activation requires influx of extracellular Ca(2+); however, the channels that mediate this influx remain unknown. Here, we tested whether the α1 subunit of the T-type channel CaV3.2, encoded by Cacna1h, mediates Ca(2+) entry into oocytes. We show that mouse eggs express a robust voltage-activated Ca(2+) current that is completely absent in Cacna1h(-/-) eggs. Cacna1h(-/-) females have reduced litter sizes, and careful analysis of Ca(2+) oscillation patterns in Cacna1h(-/-) eggs following in vitro fertilization (IVF) revealed reductions in first transient length and oscillation persistence. Total and endoplasmic reticulum (ER) Ca(2+) stores were also reduced in Cacna1h(-/-) eggs. Pharmacological inhibition of CaV3.2 in wild-type CF-1 strain eggs using mibefradil or pimozide reduced Ca(2+) store accumulation during oocyte maturation and reduced Ca(2+) oscillation persistence, frequency and number following IVF. Overall, these data show that CaV3.2 T-type channels have prev8iously unrecognized roles in supporting the meiotic-maturation-associated increase in ER Ca(2+) stores and mediating Ca(2+) influx required for the activation of development.
Keywords: Ca2+; Egg activation; Fertilization; Meiosis; Oocyte; T-type channel.
© 2015. Published by The Company of Biologists Ltd.
Conflict of interest statement
The authors declare no competing or financial interests.
Figures
References
-
- Arnoult C., Villaz M. and Florman H. M. (1998). Pharmacological properties of the T-type Ca2+ current of mouse spermatogenic cells. Mol. Pharmacol. 53, 1104-1111. - PubMed
-
- Backs J., Stein P., Backs T., Duncan F. E., Grueter C. E., McAnally J., Qi X., Schultz R. M. and Olson E. N. (2010). The gamma isoform of CaM kinase II controls mouse egg activation by regulating cell cycle resumption. Proc. Natl. Acad. Sci. USA 107, 81-86. 10.1073/pnas.0912658106 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
