

Surface Polysaccharide Mutants Reveal that Absence of O Antigen Reduces Biofilm Formation of Actinobacillus pleuropneumoniae
- PMID: 26483403
- PMCID: PMC4694004
- DOI: 10.1128/IAI.00912-15
Surface Polysaccharide Mutants Reveal that Absence of O Antigen Reduces Biofilm Formation of Actinobacillus pleuropneumoniae
Abstract
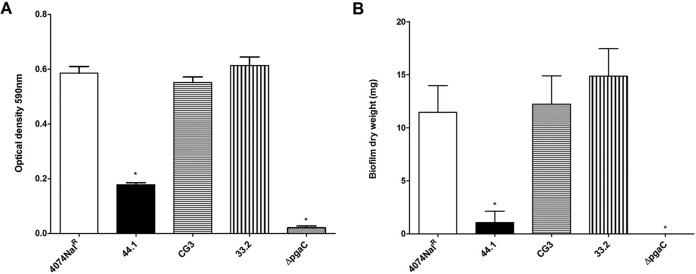
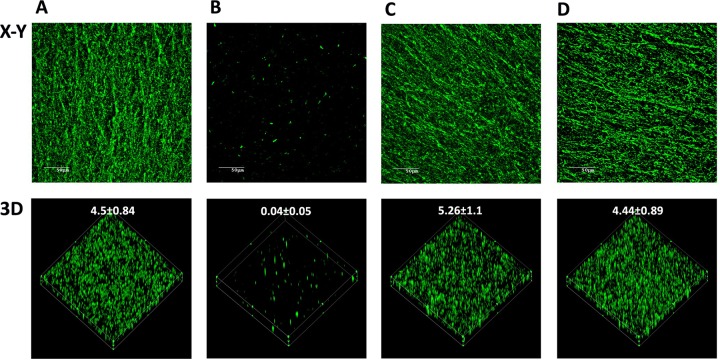
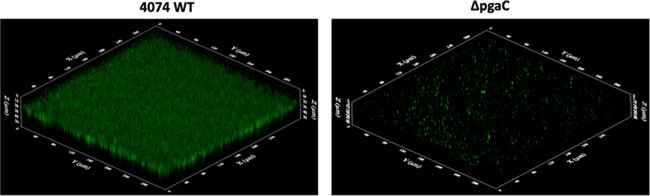
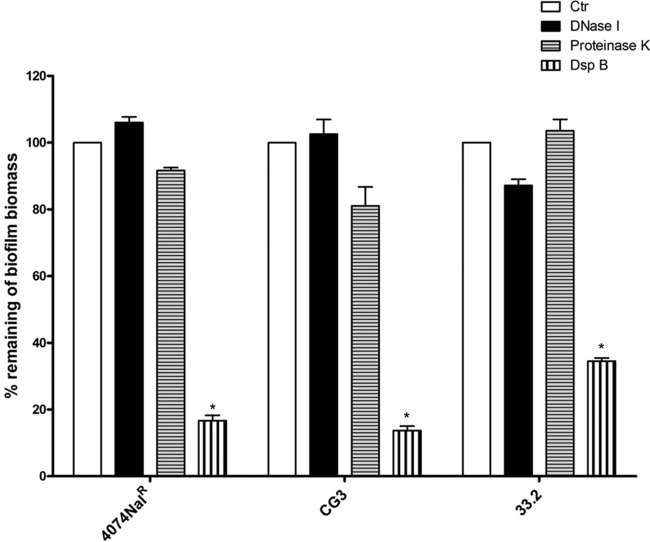
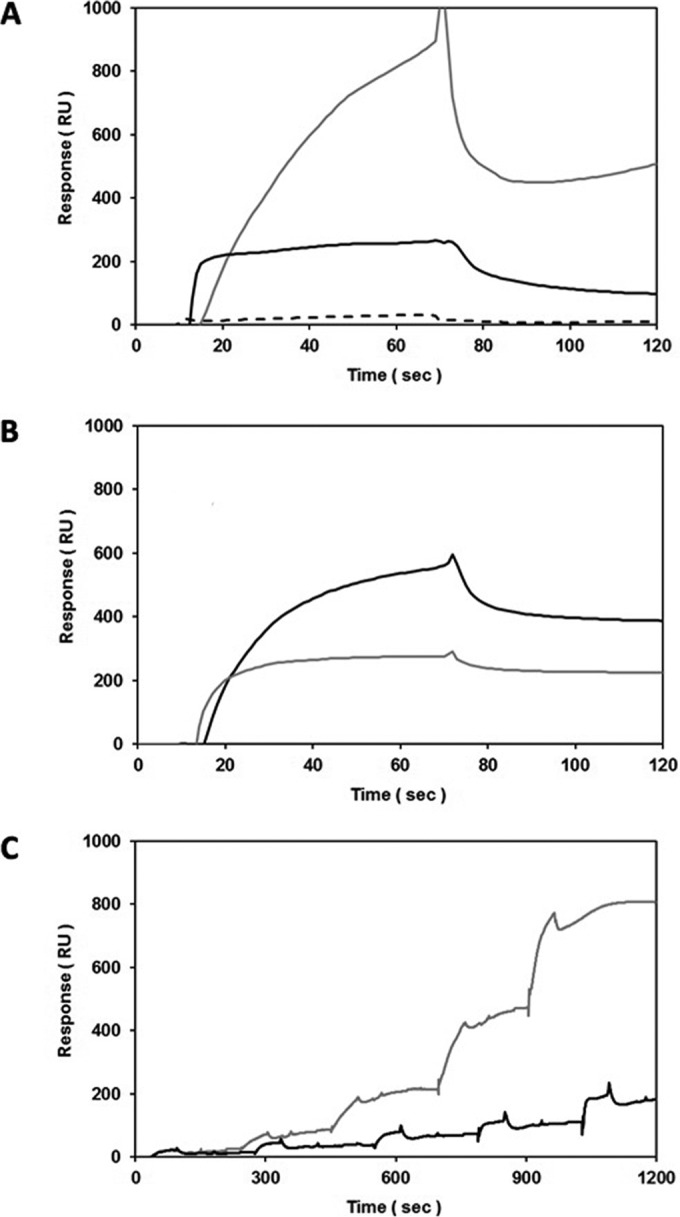
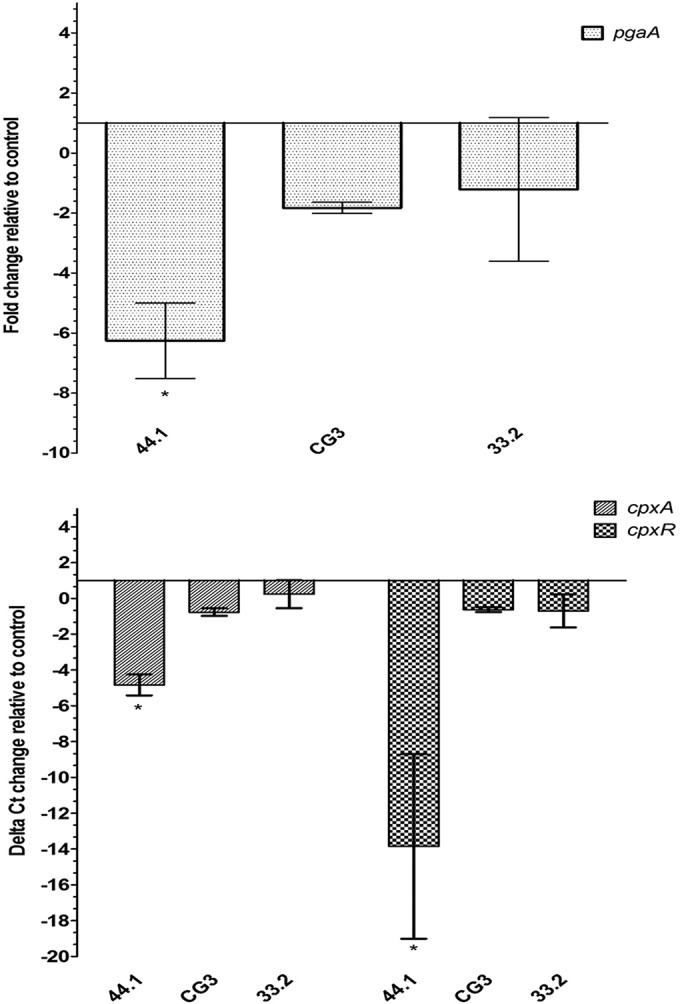
Actinobacillus pleuropneumoniae is a Gram-negative bacterium belonging to the Pasteurellaceae family and the causative agent of porcine pleuropneumonia, a highly contagious lung disease causing important economic losses. Surface polysaccharides, including lipopolysaccharides (LPS) and capsular polysaccharides (CPS), are implicated in the adhesion and virulence of A. pleuropneumoniae, but their role in biofilm formation is still unclear. In this study, we investigated the requirement for these surface polysaccharides in biofilm formation by A. pleuropneumoniae serotype 1. Well-characterized mutants were used: an O-antigen LPS mutant, a truncated core LPS mutant with an intact O antigen, a capsule mutant, and a poly-N-acetylglucosamine (PGA) mutant. We compared the amount of biofilm produced by the parental strain and the isogenic mutants using static and dynamic systems. Compared to the findings for the biofilm of the parental or other strains, the biofilm of the O antigen and the PGA mutants was dramatically reduced, and it had less cell-associated PGA. Real-time PCR analyses revealed a significant reduction in the level of pgaA, cpxR, and cpxA mRNA in the biofilm cells of the O-antigen mutant compared to that in the biofilm cells of the parental strain. Specific binding between PGA and LPS was consistently detected by surface plasmon resonance, but the lack of O antigen did not abolish these interactions. In conclusion, the absence of the O antigen reduces the ability of A. pleuropneumoniae to form a biofilm, and this is associated with the reduced expression and production of PGA.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures

References
-
- Gottschalk M. 2012. Actinobacillosis, p 653–669. In Karriker L, Ramirez A, Schwartz K, Stevenson G, Zimmerman J (ed), Diseases of swine, 10th ed John Wiley & Sons, Inc, Hoboken, NJ.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
