

Topographic representations of object size and relationships with numerosity reveal generalized quantity processing in human parietal cortex
- PMID: 26483452
- PMCID: PMC4640722
- DOI: 10.1073/pnas.1515414112
Topographic representations of object size and relationships with numerosity reveal generalized quantity processing in human parietal cortex
Abstract
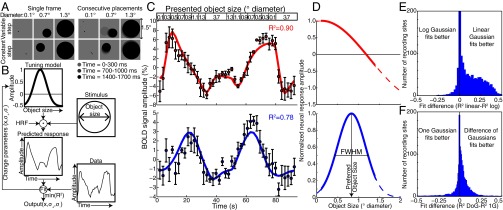
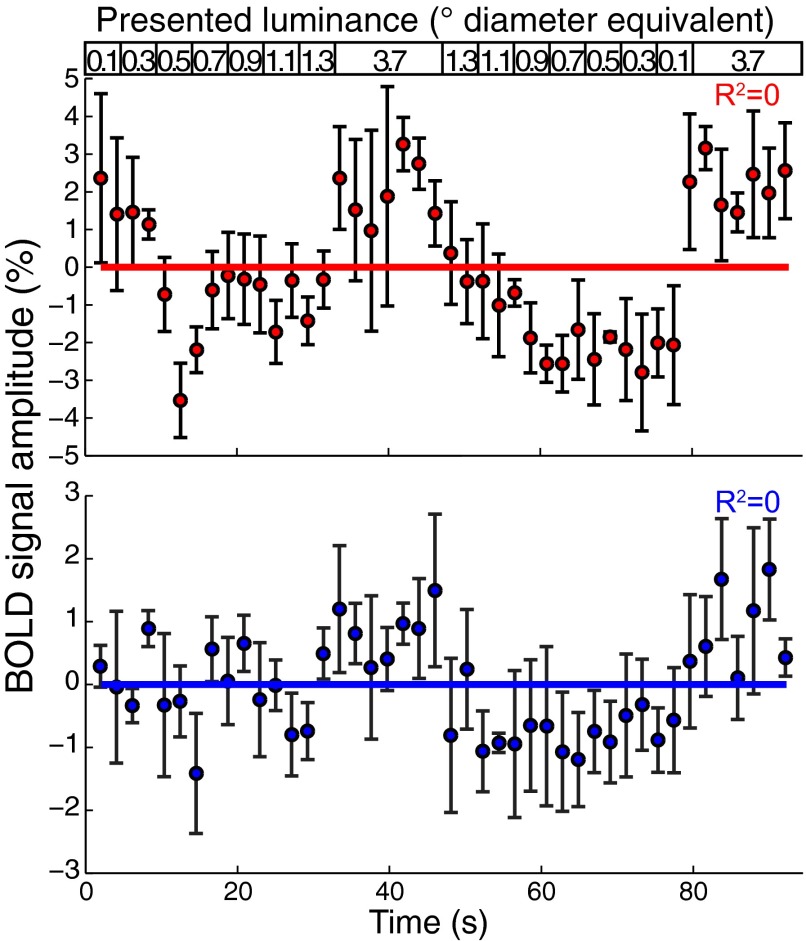
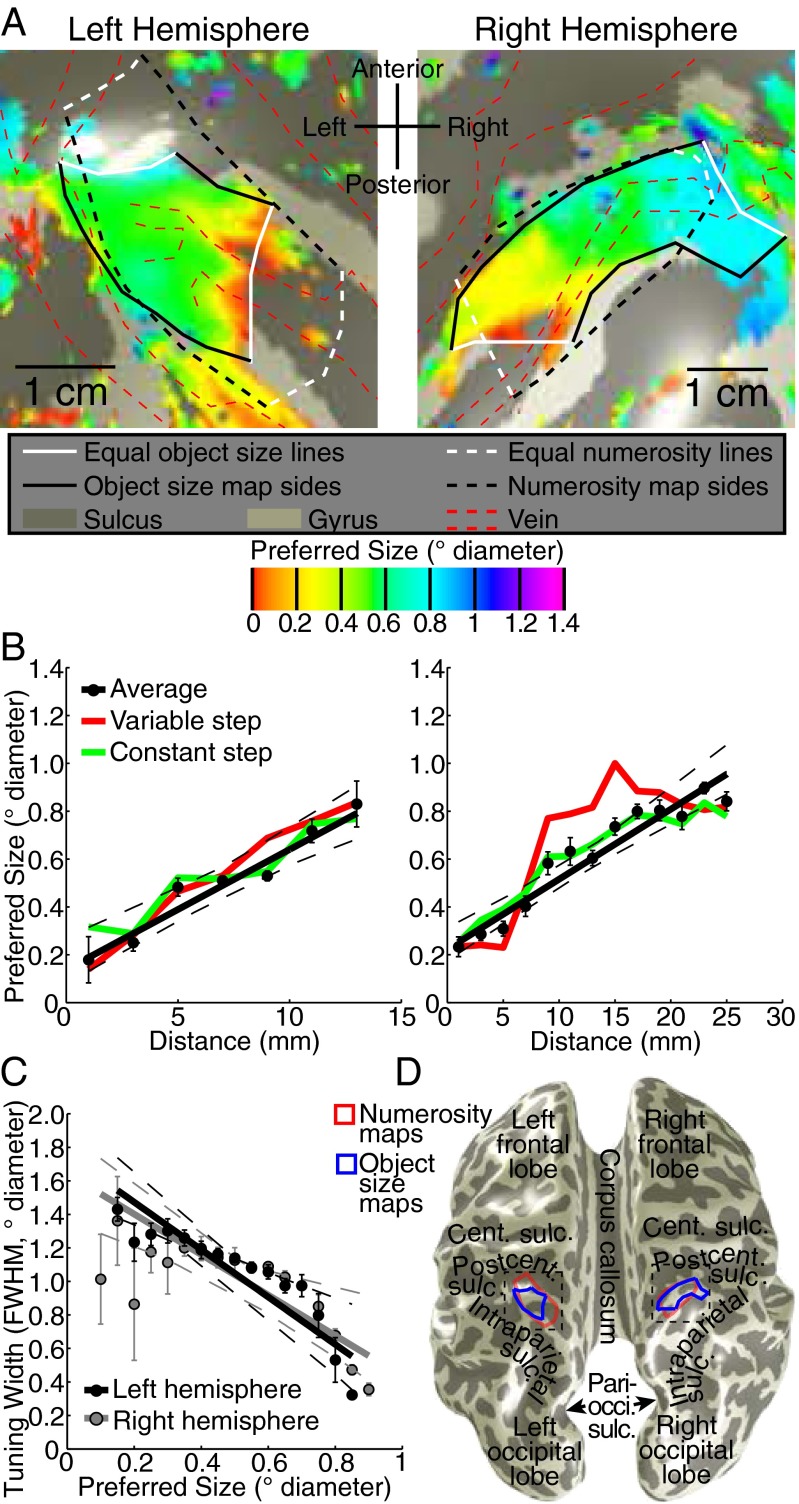
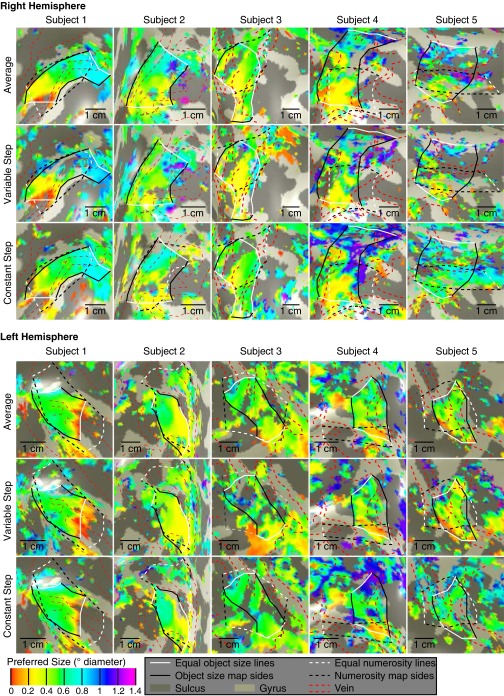
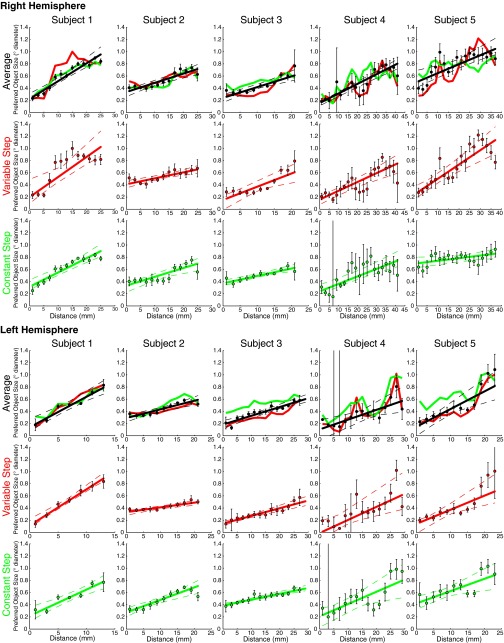
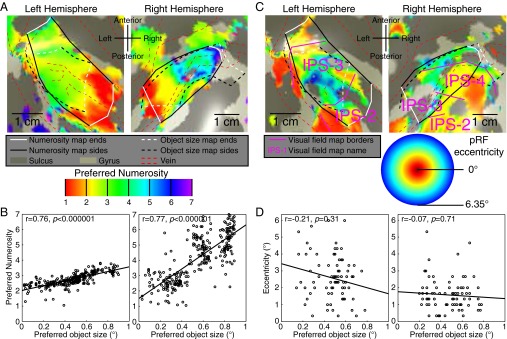
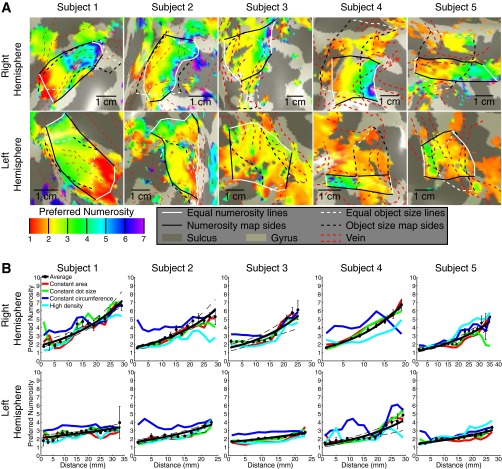
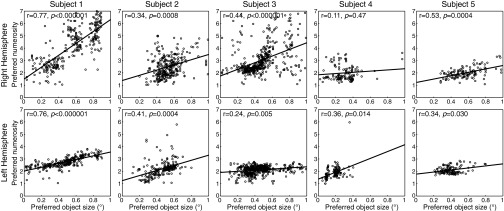
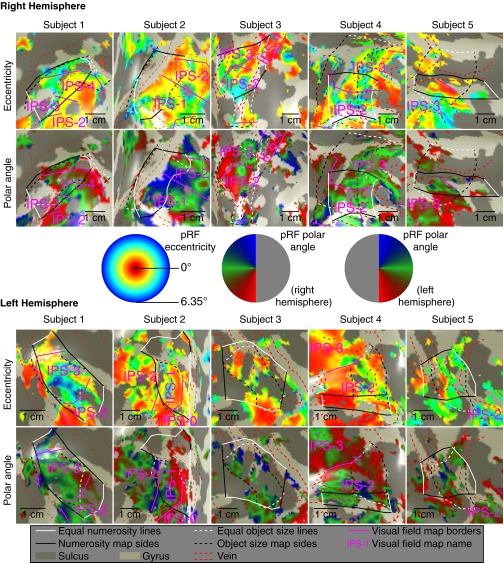
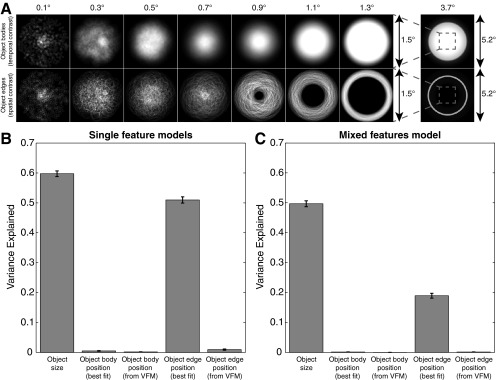
Humans and many animals analyze sensory information to estimate quantities that guide behavior and decisions. These quantities include numerosity (object number) and object size. Having recently demonstrated topographic maps of numerosity, we ask whether the brain also contains maps of object size. Using ultra-high-field (7T) functional MRI and population receptive field modeling, we describe tuned responses to visual object size in bilateral human posterior parietal cortex. Tuning follows linear Gaussian functions and shows surround suppression, and tuning width narrows with increasing preferred object size. Object size-tuned responses are organized in bilateral topographic maps, with similar cortical extents responding to large and small objects. These properties of object size tuning and map organization all differ from the numerosity representation, suggesting that object size and numerosity tuning result from distinct mechanisms. However, their maps largely overlap and object size preferences correlate with numerosity preferences, suggesting associated representations of these two quantities. Object size preferences here show no discernable relation to visual position preferences found in visuospatial receptive fields. As such, object size maps (much like numerosity maps) do not reflect sensory organ structure but instead emerge within the brain. We speculate that, as in sensory processing, optimization of cognitive processing using topographic maps may be a common organizing principle in association cortex. Interactions between object size and numerosity maps may associate cognitive representations of these related features, potentially allowing consideration of both quantities together when making decisions.
Keywords: high-field 7T fMRI; numerosity; object size; topographic maps.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





Similar articles
-
Topographic representation of numerosity in the human parietal cortex.Science. 2013 Sep 6;341(6150):1123-6. doi: 10.1126/science.1239052. Science. 2013. PMID: 24009396
-
Cortical quantity representations of visual numerosity and timing overlap increasingly into superior cortices but remain distinct.Neuroimage. 2024 Feb 1;286:120515. doi: 10.1016/j.neuroimage.2024.120515. Epub 2024 Jan 10. Neuroimage. 2024. PMID: 38216105
-
Size constancy affects the perception and parietal neural representation of object size.Neuroimage. 2021 May 15;232:117909. doi: 10.1016/j.neuroimage.2021.117909. Epub 2021 Feb 27. Neuroimage. 2021. PMID: 33652148
-
The role of neural tuning in quantity perception.Trends Cogn Sci. 2022 Jan;26(1):11-24. doi: 10.1016/j.tics.2021.10.004. Epub 2021 Oct 23. Trends Cogn Sci. 2022. PMID: 34702662 Review.
-
Comparing Parietal Quantity-Processing Mechanisms between Humans and Macaques.Trends Cogn Sci. 2017 Oct;21(10):779-793. doi: 10.1016/j.tics.2017.07.002. Epub 2017 Aug 9. Trends Cogn Sci. 2017. PMID: 28802806 Review.
Cited by
-
Inferior Occipital Gyrus Is Organized along Common Gradients of Spatial and Face-Part Selectivity.J Neurosci. 2021 Jun 23;41(25):5511-5521. doi: 10.1523/JNEUROSCI.2415-20.2021. Epub 2021 May 20. J Neurosci. 2021. PMID: 34016715 Free PMC article.
-
A MEG Study on the Processing of Time and Quantity: Parietal Overlap but Functional Divergence.Front Psychol. 2019 Feb 4;10:139. doi: 10.3389/fpsyg.2019.00139. eCollection 2019. Front Psychol. 2019. PMID: 30778314 Free PMC article.
-
View Normalization of Object Size in the Right Parietal Cortex.Vision (Basel). 2022 Jul 1;6(3):41. doi: 10.3390/vision6030041. Vision (Basel). 2022. PMID: 35893758 Free PMC article.
-
How is visual separation assessed? By counting distance units.Front Psychol. 2024 May 30;15:1410297. doi: 10.3389/fpsyg.2024.1410297. eCollection 2024. Front Psychol. 2024. PMID: 38873519 Free PMC article.
-
Position- and scale-invariant object-centered spatial localization in monkey frontoparietal cortex dynamically adapts to cognitive demand.Nat Commun. 2024 Apr 18;15(1):3357. doi: 10.1038/s41467-024-47554-4. Nat Commun. 2024. PMID: 38637493 Free PMC article.
References
-
- Woodruff G, Premack D, Kennel K. Conservation of liquid and solid quantity by the chimpanzee. Science. 1978;202(4371):991–994. - PubMed
-
- Dehaene S, Spelke E, Pinel P, Stanescu R, Tsivkin S. Sources of mathematical thinking: Behavioral and brain-imaging evidence. Science. 1999;284(5416):970–974. - PubMed
-
- Burr D, Ross J. A visual sense of number. Curr Biol. 2008;18(6):425–428. - PubMed
-
- Walsh V. A theory of magnitude: Common cortical metrics of time, space and quantity. Trends Cogn Sci. 2003;7(11):483–488. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
