

Synaptic circuits and their variations within different columns in the visual system of Drosophila
- PMID: 26483464
- PMCID: PMC4640747
- DOI: 10.1073/pnas.1509820112
Synaptic circuits and their variations within different columns in the visual system of Drosophila
Abstract
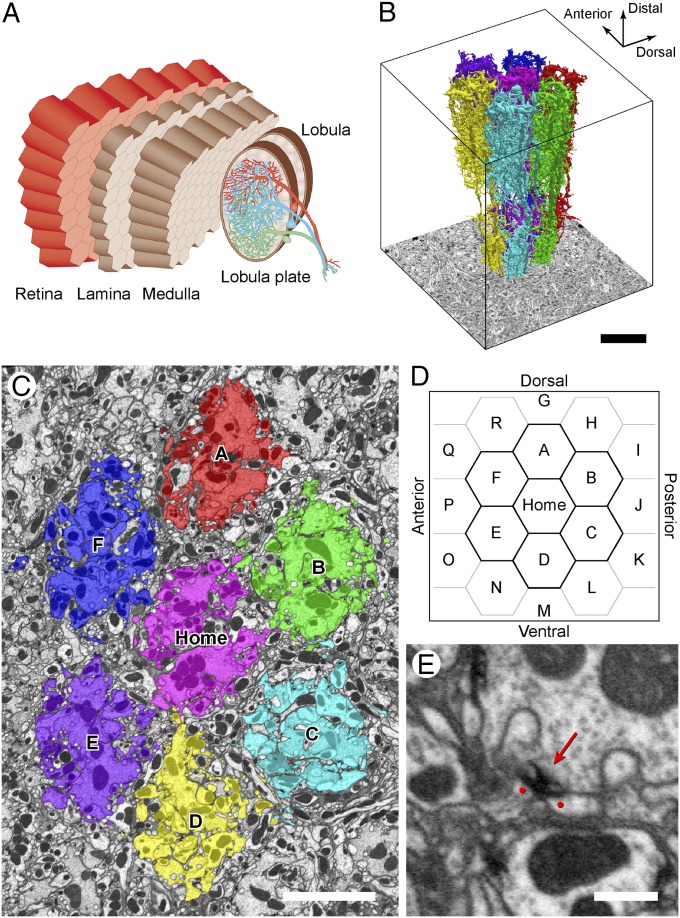
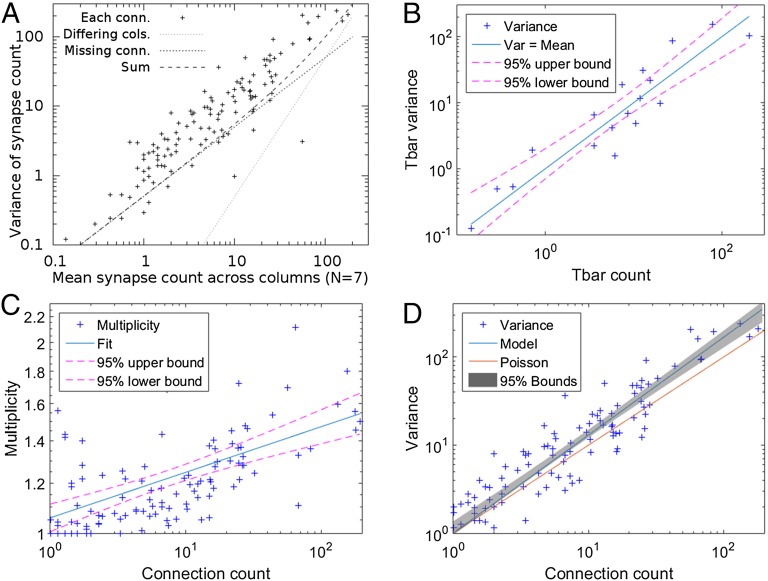
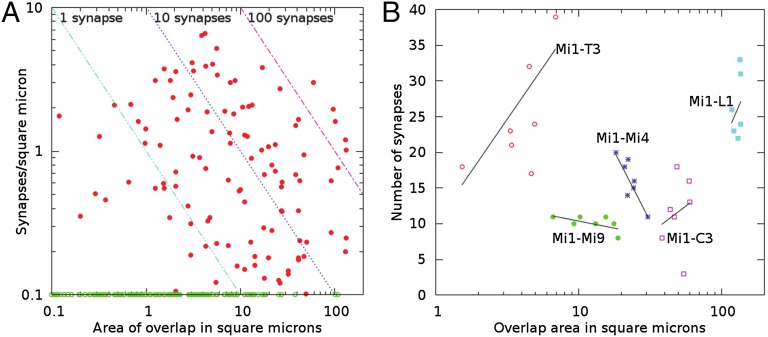
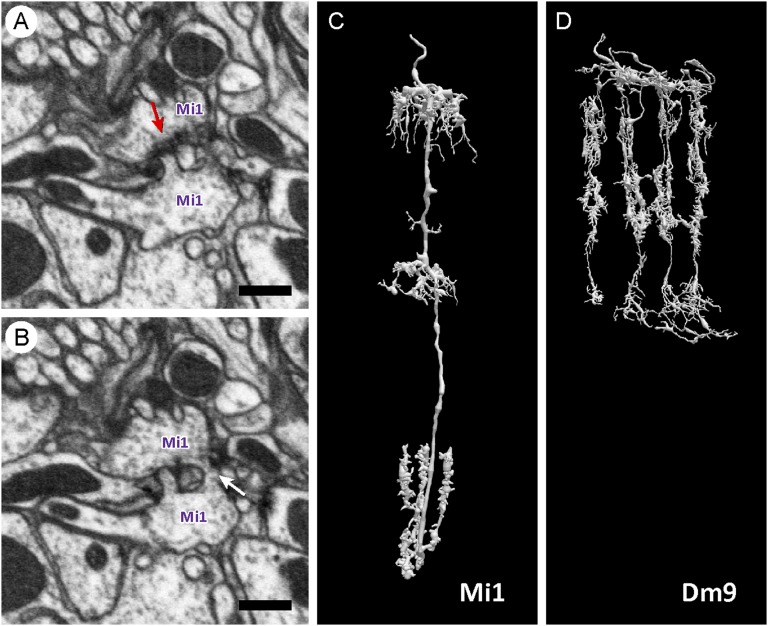
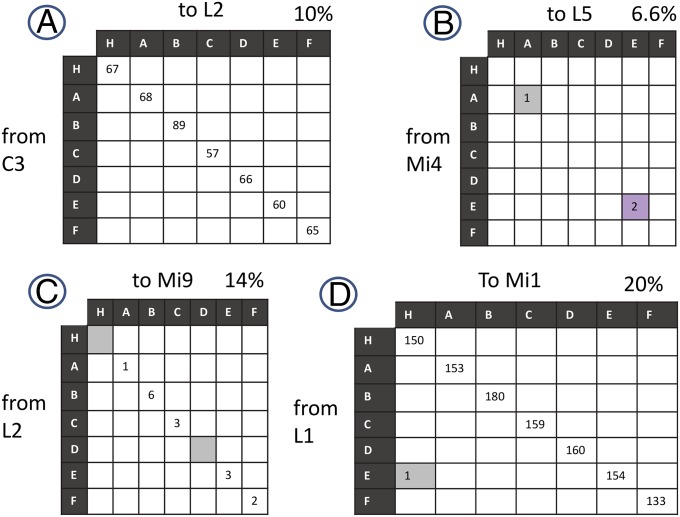
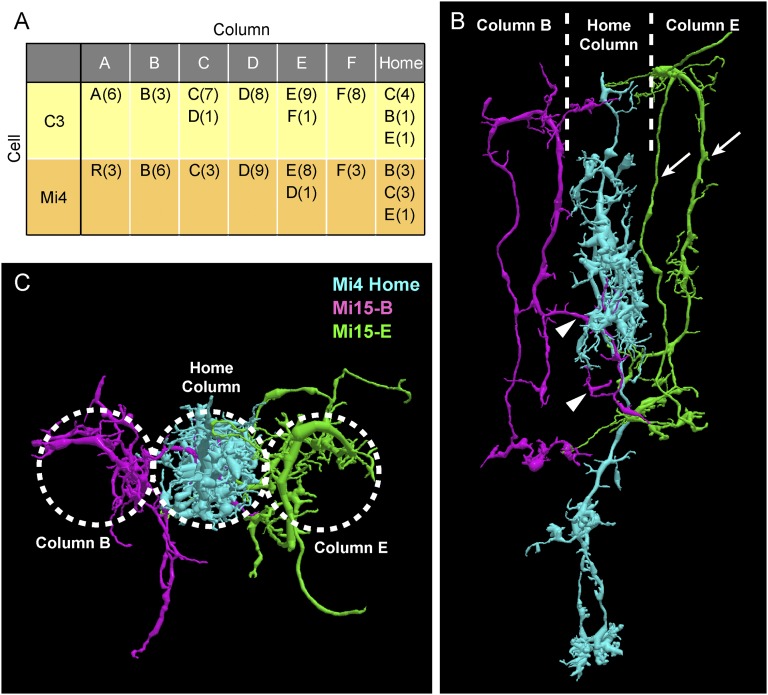
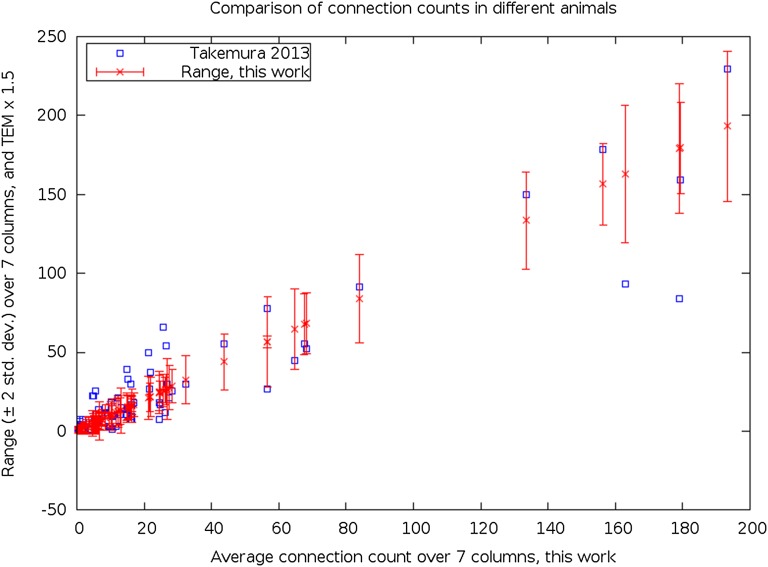
We reconstructed the synaptic circuits of seven columns in the second neuropil or medulla behind the fly's compound eye. These neurons embody some of the most stereotyped circuits in one of the most miniaturized of animal brains. The reconstructions allow us, for the first time to our knowledge, to study variations between circuits in the medulla's neighboring columns. This variation in the number of synapses and the types of their synaptic partners has previously been little addressed because methods that visualize multiple circuits have not resolved detailed connections, and existing connectomic studies, which can see such connections, have not so far examined multiple reconstructions of the same circuit. Here, we address the omission by comparing the circuits common to all seven columns to assess variation in their connection strengths and the resultant rates of several different and distinct types of connection error. Error rates reveal that, overall, <1% of contacts are not part of a consensus circuit, and we classify those contacts that supplement (E+) or are missing from it (E-). Autapses, in which the same cell is both presynaptic and postsynaptic at the same synapse, are occasionally seen; two cells in particular, Dm9 and Mi1, form ≥ 20-fold more autapses than do other neurons. These results delimit the accuracy of developmental events that establish and normally maintain synaptic circuits with such precision, and thereby address the operation of such circuits. They also establish a precedent for error rates that will be required in the new science of connectomics.
Keywords: biological error rates; neural circuits; reconstruction error rates; stereotypy.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Chklovskii DB, Schikorski T, Stevens CF. Wiring optimization in cortical circuits. Neuron. 2002;34(3):341–347. - PubMed
-
- Cherniak C. Neural wiring optimization. Prog Brain Res. 2012;195:361–371. - PubMed
-
- Robertson RM. Neuronal circuits controlling flight in the locust: Central generation of the rhythm. Trends Neurosci. 1986;9(6):278–280.
-
- Grillner S, Wallén P. Cellular bases of a vertebrate locomotor system-steering, intersegmental and segmental co-ordination and sensory control. Brain Res Brain Res Rev. 2002;40(1-3):92–106. - PubMed
-
- Kristan WB, Jr, Calabrese RL, Friesen WO. Neuronal control of leech behavior. Prog Neurobiol. 2005;76(5):279–327. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
