

Coevolutionary patterning of teeth and taste buds
- PMID: 26483492
- PMCID: PMC4640805
- DOI: 10.1073/pnas.1514298112
Coevolutionary patterning of teeth and taste buds
Abstract
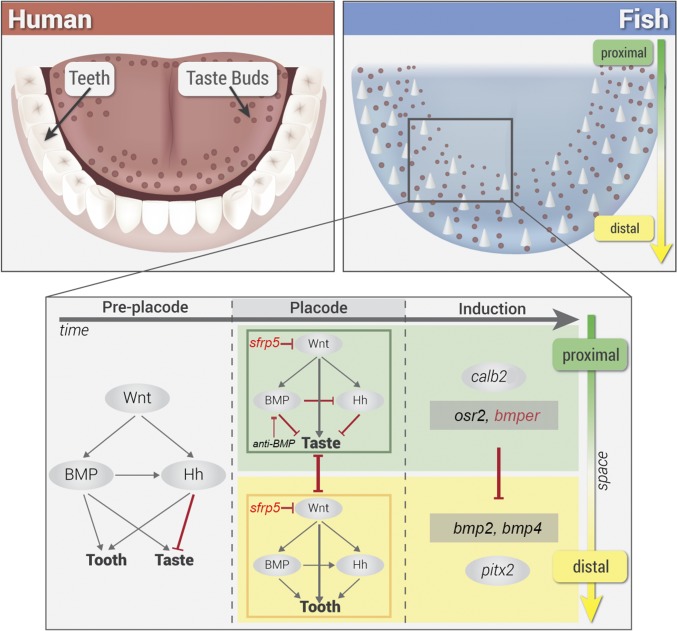
Teeth and taste buds are iteratively patterned structures that line the oro-pharynx of vertebrates. Biologists do not fully understand how teeth and taste buds develop from undifferentiated epithelium or how variation in organ density is regulated. These organs are typically studied independently because of their separate anatomical location in mammals: teeth on the jaw margin and taste buds on the tongue. However, in many aquatic animals like bony fishes, teeth and taste buds are colocalized one next to the other. Using genetic mapping in cichlid fishes, we identified shared loci controlling a positive correlation between tooth and taste bud densities. Genome intervals contained candidate genes expressed in tooth and taste bud fields. sfrp5 and bmper, notable for roles in Wingless (Wnt) and bone morphogenetic protein (BMP) signaling, were differentially expressed across cichlid species with divergent tooth and taste bud density, and were expressed in the development of both organs in mice. Synexpression analysis and chemical manipulation of Wnt, BMP, and Hedgehog (Hh) pathways suggest that a common cichlid oral lamina is competent to form teeth or taste buds. Wnt signaling couples tooth and taste bud density and BMP and Hh mediate distinct organ identity. Synthesizing data from fish and mouse, we suggest that the Wnt-BMP-Hh regulatory hierarchy that configures teeth and taste buds on mammalian jaws and tongues may be an evolutionary remnant inherited from ancestors wherein these organs were copatterned from common epithelium.
Keywords: bipotency; placode patterning; plasticity; quantitative trait loci; tooth/taste bud development.
Conflict of interest statement
The authors declare no conflict of interest.
Figures






Similar articles
-
Developmental Plasticity of Patterned and Regenerating Oral Organs.Curr Top Dev Biol. 2015;115:321-33. doi: 10.1016/bs.ctdb.2015.07.005. Epub 2015 Oct 6. Curr Top Dev Biol. 2015. PMID: 26589931 Free PMC article. Review.
-
Developmental plasticity of epithelial stem cells in tooth and taste bud renewal.Proc Natl Acad Sci U S A. 2019 Sep 3;116(36):17858-17866. doi: 10.1073/pnas.1821202116. Epub 2019 Aug 19. Proc Natl Acad Sci U S A. 2019. PMID: 31427537 Free PMC article.
-
Multiple Shh signaling centers participate in fungiform papilla and taste bud formation and maintenance.Dev Biol. 2013 Oct 1;382(1):82-97. doi: 10.1016/j.ydbio.2013.07.022. Epub 2013 Aug 2. Dev Biol. 2013. PMID: 23916850 Free PMC article.
-
Pleiotropic functions of embryonic sonic hedgehog expression link jaw and taste bud amplification with eye loss during cavefish evolution.Dev Biol. 2009 Jun 1;330(1):200-11. doi: 10.1016/j.ydbio.2009.03.003. Epub 2009 Mar 11. Dev Biol. 2009. PMID: 19285488 Free PMC article.
-
Hedgehog Signaling Regulates Taste Organs and Oral Sensation: Distinctive Roles in the Epithelium, Stroma, and Innervation.Int J Mol Sci. 2019 Mar 16;20(6):1341. doi: 10.3390/ijms20061341. Int J Mol Sci. 2019. PMID: 30884865 Free PMC article. Review.
Cited by
-
The metamorphic transition of the frog mouth: from tadpole keratinized mouthparts to adult teeth.R Soc Open Sci. 2025 Sep 3;12(9):251196. doi: 10.1098/rsos.251196. eCollection 2025 Sep. R Soc Open Sci. 2025. PMID: 40904991 Free PMC article.
-
Genetic analyses in Lake Malawi cichlids identify new roles for Fgf signaling in scale shape variation.Commun Biol. 2018 May 31;1:55. doi: 10.1038/s42003-018-0060-4. eCollection 2018. Commun Biol. 2018. PMID: 30271938 Free PMC article.
-
Identification of prohormones and pituitary neuropeptides in the African cichlid, Astatotilapia burtoni.BMC Genomics. 2016 Aug 19;17(1):660. doi: 10.1186/s12864-016-2914-9. BMC Genomics. 2016. PMID: 27543050 Free PMC article.
-
Meristic co-evolution and genomic co-localization of lateral line scales and vertebrae in Central American cichlid fishes.Ecol Evol. 2024 Sep 15;14(9):e70266. doi: 10.1002/ece3.70266. eCollection 2024 Sep. Ecol Evol. 2024. PMID: 39279804 Free PMC article.
-
Nanoscale and Macroscale Scaffolds with Controlled-Release Polymeric Systems for Dental Craniomaxillofacial Tissue Engineering.Materials (Basel). 2018 Aug 20;11(8):1478. doi: 10.3390/ma11081478. Materials (Basel). 2018. PMID: 30127246 Free PMC article. Review.
References
-
- Tucker A, Sharpe P. The cutting-edge of mammalian development; how the embryo makes teeth. Nat Rev Genet. 2004;5(7):499–508. - PubMed
-
- Kondo S, Miura T. Reaction-diffusion model as a framework for understanding biological pattern formation. Science. 2010;329(5999):1616–1620. - PubMed
-
- Sick S, Reinker S, Timmer J, Schlake T. WNT and DKK determine hair follicle spacing through a reaction-diffusion mechanism. Science. 2006;314(5804):1447–1450. - PubMed
Publication types
MeSH terms
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
