

Peroxisomal Import Reduces the Proapoptotic Activity of Deubiquitinating Enzyme USP2
- PMID: 26484888
- PMCID: PMC4617714
- DOI: 10.1371/journal.pone.0140685
Peroxisomal Import Reduces the Proapoptotic Activity of Deubiquitinating Enzyme USP2
Abstract
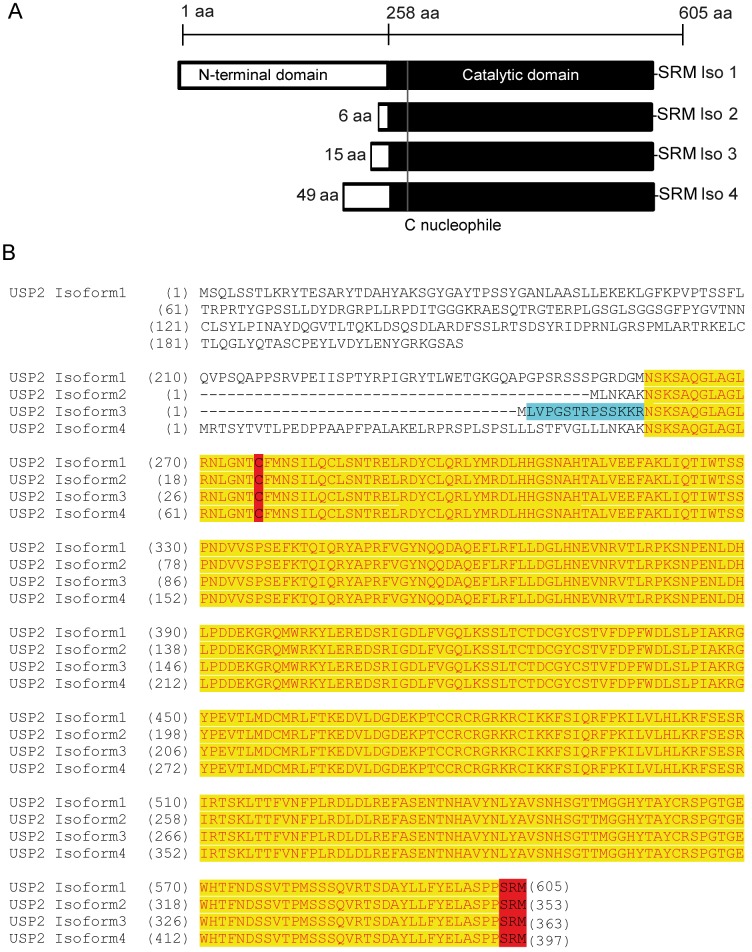
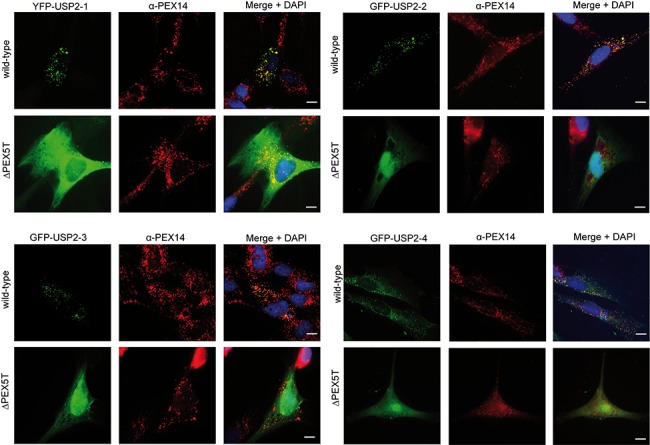
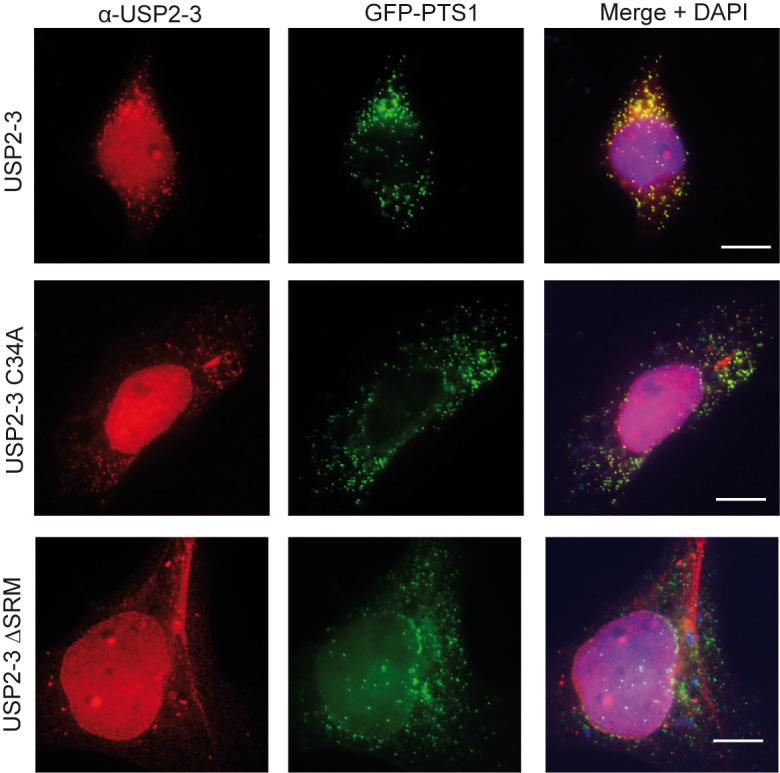
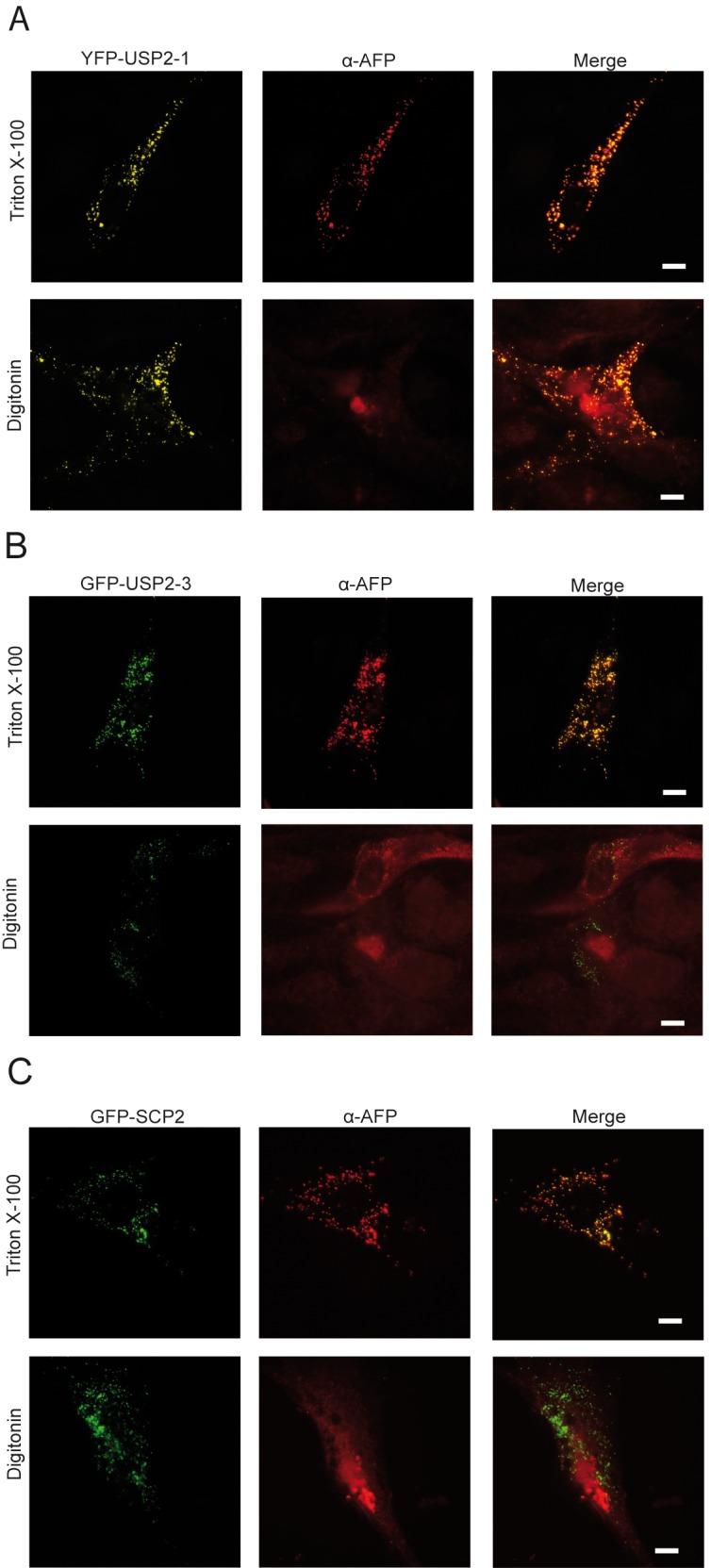
The human deubiquitinating enzyme ubiquitin-specific protease 2 (USP2) regulates multiple cellular pathways, including cell proliferation and apoptosis. As a result of alternative splicing four USP2 isoenzymes are expressed in human cells of which all contain a weak peroxisome targeting signal of type 1 (PTS1) at their C-termini. Here, we systematically analyzed apoptotic effects induced by overexpression and intracellular localization for each isoform. All isoforms exhibit proapoptotic activity and are post-translationally imported into the matrix of peroxisomes in a PEX5-dependent manner. However, a significant fraction of the USP2 pool resides in the cytosol due to a weaker PTS1 and thus low affinity to the PTS receptor PEX5. Blocking of peroxisomal import did not interfere with the proapoptotic activity of USP2, suggesting that the enzyme performs its critical function outside of this compartment. Instead, increase of the efficiency of USP2 import into peroxisomes either by optimization of its peroxisomal targeting signal or by overexpression of the PTS1 receptor did result in a reduction of the apoptotic rate of transfected cells. Our studies suggest that peroxisomal import of USP2 provides additional control over the proapoptotic activity of cytosolic USP2 by spatial separation of the deubiquitinating enzymes from their interaction partners in the cytosol and nucleus.
Conflict of interest statement
Figures
Similar articles
-
Peroxisomal ubiquitin-protein ligases peroxin2 and peroxin10 have distinct but synergistic roles in matrix protein import and peroxin5 retrotranslocation in Arabidopsis.Plant Physiol. 2014 Nov;166(3):1329-44. doi: 10.1104/pp.114.247148. Epub 2014 Sep 11. Plant Physiol. 2014. PMID: 25214533 Free PMC article.
-
The peroxisomal import receptor PEX5 functions as a stress sensor, retaining catalase in the cytosol in times of oxidative stress.Biochim Biophys Acta Mol Cell Res. 2017 Oct;1864(10):1833-1843. doi: 10.1016/j.bbamcr.2017.07.013. Epub 2017 Jul 29. Biochim Biophys Acta Mol Cell Res. 2017. PMID: 28760655
-
PEX5, the shuttling import receptor for peroxisomal matrix proteins, is a redox-sensitive protein.Traffic. 2014 Jan;15(1):94-103. doi: 10.1111/tra.12129. Epub 2013 Oct 31. Traffic. 2014. PMID: 24118911
-
Towards solving the mystery of peroxisomal matrix protein import.Trends Cell Biol. 2024 May;34(5):388-405. doi: 10.1016/j.tcb.2023.08.005. Epub 2023 Sep 22. Trends Cell Biol. 2024. PMID: 37743160 Free PMC article. Review.
-
Peroxisomal matrix protein receptor ubiquitination and recycling.Biochim Biophys Acta. 2006 Dec;1763(12):1620-8. doi: 10.1016/j.bbamcr.2006.08.046. Epub 2006 Sep 3. Biochim Biophys Acta. 2006. PMID: 17028012 Review.
Cited by
-
Diffusion and interaction dynamics of the cytosolic peroxisomal import receptor PEX5.Biophys Rep (N Y). 2022 Jun 8;2(2):None. doi: 10.1016/j.bpr.2022.100055. Biophys Rep (N Y). 2022. PMID: 36299769 Free PMC article.
-
The Molecular Mechanisms of Regulation on USP2's Alternative Splicing and the Significance of Its Products.Int J Biol Sci. 2017 Nov 2;13(12):1489-1496. doi: 10.7150/ijbs.21637. eCollection 2017. Int J Biol Sci. 2017. PMID: 29230097 Free PMC article. Review.
-
USP2-Related Cellular Signaling and Consequent Pathophysiological Outcomes.Int J Mol Sci. 2021 Jan 26;22(3):1209. doi: 10.3390/ijms22031209. Int J Mol Sci. 2021. PMID: 33530560 Free PMC article. Review.
-
Dysregulation and activities of ubiquitin specific peptidase 2b in the pathogenesis of hepatocellular carcinoma.Am J Cancer Res. 2021 Oct 15;11(10):4746-4767. eCollection 2021. Am J Cancer Res. 2021. PMID: 34765291 Free PMC article.
-
Systematic multi-level analysis of an organelle proteome reveals new peroxisomal functions.Mol Syst Biol. 2022 Sep;18(9):e11186. doi: 10.15252/msb.202211186. Mol Syst Biol. 2022. PMID: 36164978 Free PMC article.
References
-
- Chau V, Tobias JW, Bachmair A, Marriott D, Ecker DJ, Gonda DK, et al. A multiubiquitin chain is confined to specific lysine in a targeted short-lived protein. Science. 1989;243:1576–83. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
