

Host and microbiota interactions are critical for development of murine Crohn's-like ileitis
- PMID: 26487367
- PMCID: PMC5027991
- DOI: 10.1038/mi.2015.102
Host and microbiota interactions are critical for development of murine Crohn's-like ileitis
Abstract
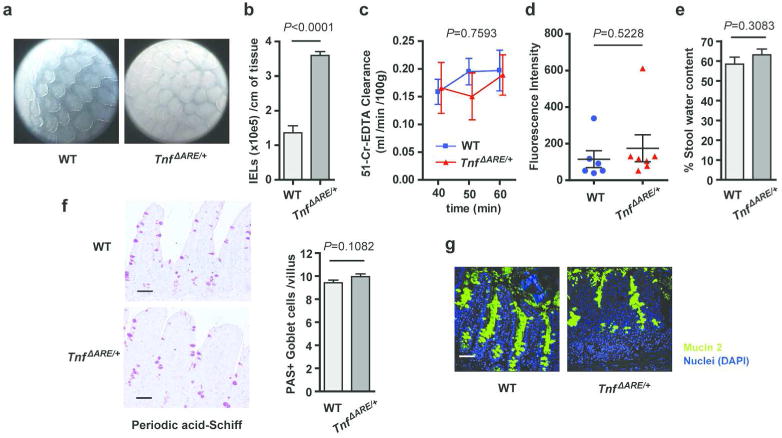
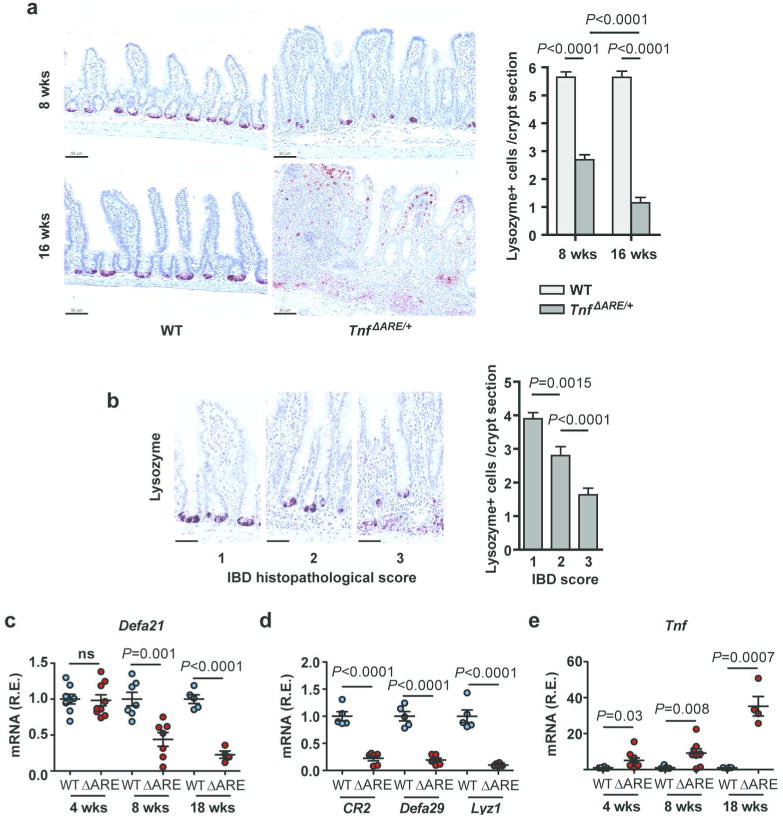
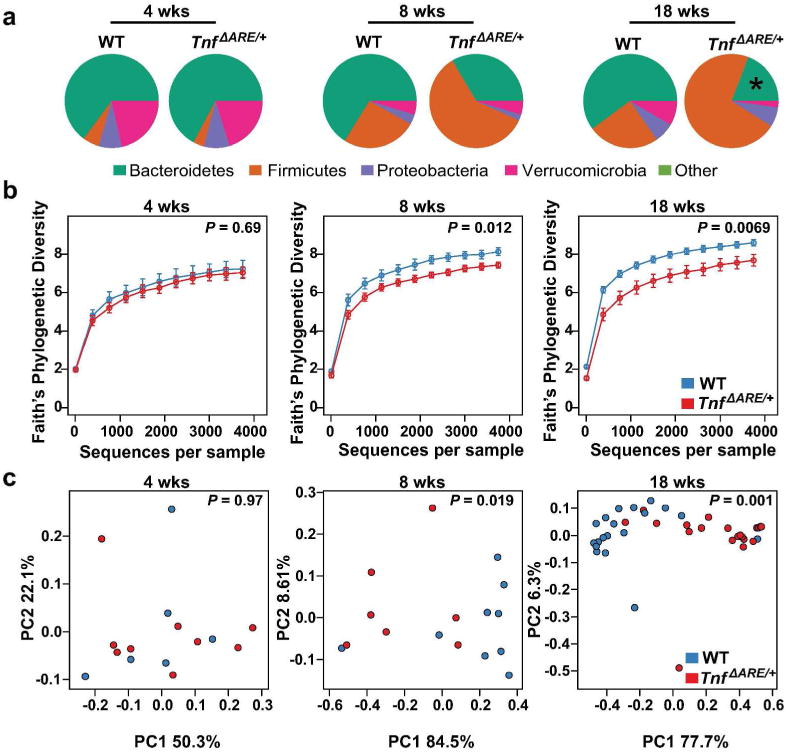
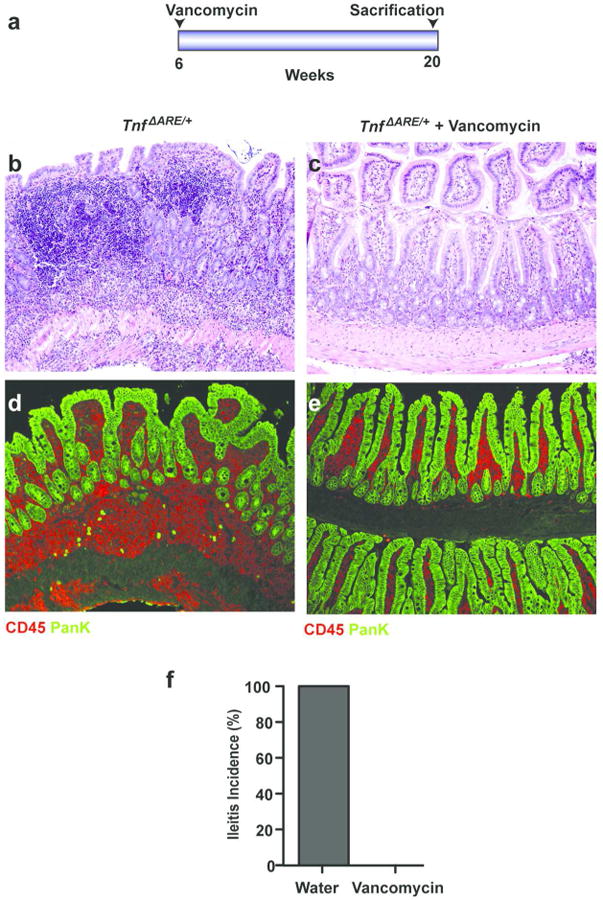
Deregulation of host-microbiota interactions in the gut is a pivotal characteristic of Crohn's disease. It remains unclear, however, whether commensals and/or the dysbiotic microbiota associated with pathology in humans are causally involved in Crohn's pathogenesis. Here, we show that Crohn's-like ileitis in Tnf(ΔARE/+) mice is microbiota-dependent. Germ-free Tnf(ΔARE/+) mice are disease-free and the microbiota and its innate recognition through Myd88 are indispensable for tumor necrosis factor (TNF) overexpression and disease initiation in this model. The epithelium of diseased mice shows no major defects in mucus barrier and paracellular permeability. However, Tnf(ΔARE/+) ileitis associates with the reduction of lysozyme-expressing Paneth cells, mediated by adaptive immune effectors. Furthermore, we show that established but not early ileitis in Tnf(ΔARE/+) mice involves defective expression of antimicrobials and dysbiosis, characterized by Firmicutes expansion, including epithelial-attaching segmented filamentous bacteria, and decreased abundance of Bacteroidetes. Microbiota modulation by antibiotic treatment at an early disease stage rescues ileitis. Our results suggest that the indigenous microbiota is sufficient to drive TNF overexpression and Crohn's ileitis in the genetically susceptible Tnf(ΔARE/+) hosts, whereas dysbiosis in this model results from disease-associated alterations including loss of lysozyme-expressing Paneth cells.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures




References
-
- Xavier RJ, Podolsky DK. Unravelling the pathogenesis of inflammatory bowel disease. Nature. 2007;448(7152):427–434. - PubMed
-
- Bevins CL, Salzman NH. Paneth cells, antimicrobial peptides and maintenance of intestinal homeostasis. Nat Rev Microbiol. 2011;9(5):356–368. - PubMed
-
- Pearson C, Uhlig HH, Powrie F. Lymphoid microenvironments and innate lymphoid cells in the gut. Trends in immunology. 2012;33(6):289–296. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
