

Protein Arginine Methyltransferase 6 (Prmt6) Is Essential for Early Zebrafish Development through the Direct Suppression of gadd45αa Stress Sensor Gene
- PMID: 26487724
- PMCID: PMC4697175
- DOI: 10.1074/jbc.M115.666347
Protein Arginine Methyltransferase 6 (Prmt6) Is Essential for Early Zebrafish Development through the Direct Suppression of gadd45αa Stress Sensor Gene
Abstract
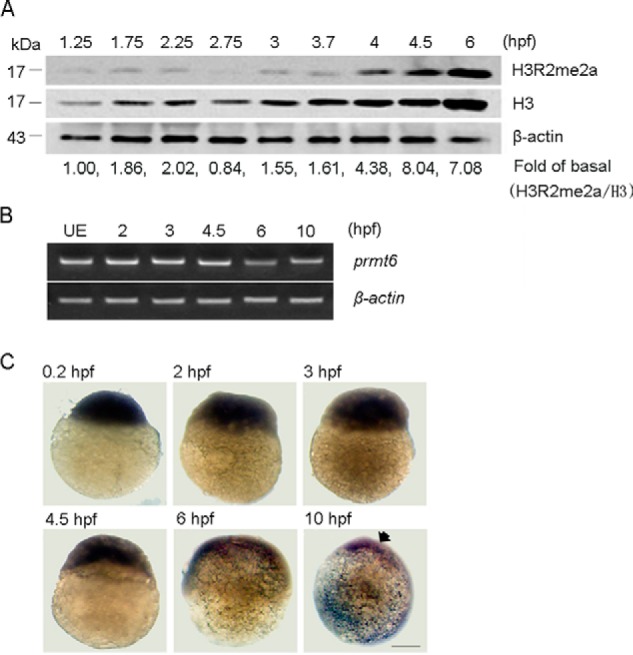
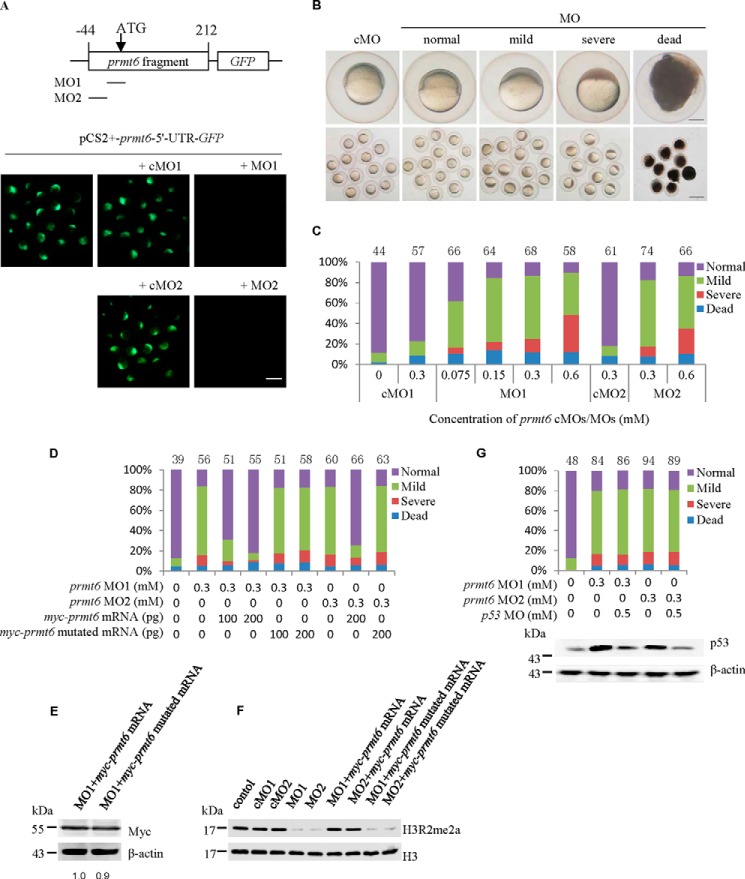
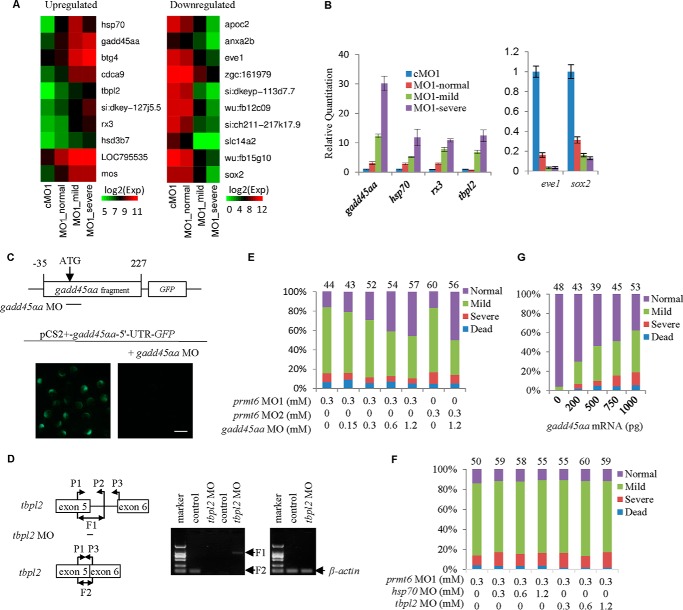
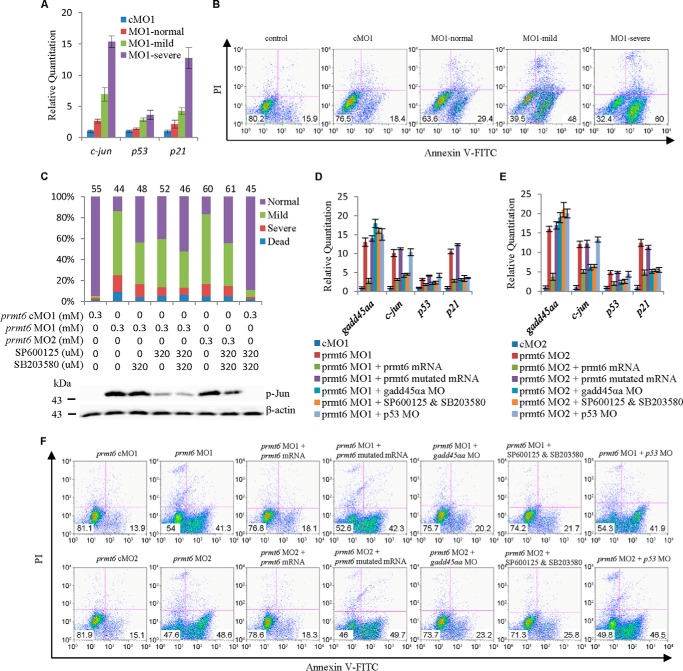
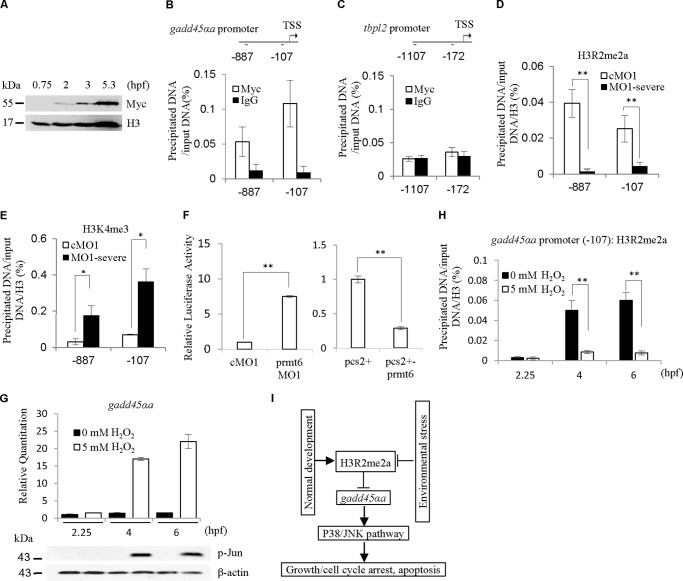
Histone lysine methylation is important in early zebrafish development; however, the role of histone arginine methylation in this process remains unclear. H3R2me2a, generated by protein arginine methyltransferase 6 (Prmt6), is a repressive mark. To explore the role of Prmt6 and H3R2me2a during zebrafish embryogenesis, we identified the maternal characteristic of prmt6 and designed two prmt6-specific morpholino-oligos (MOs) to study its importance in early development, application of which led to early epiboly defects and significantly reduced the level of H3R2me2a marks. prmt6 mRNA could rescue the epiboly defects and the H3R2me2a reduction in the prmt6 morphants. Functionally, microarray data demonstrated that growth arrest and DNA damage-inducible, α, a (gadd45αa) was a significantly up-regulated gene in MO-treated embryos, the activity of which was linked to the activation of the p38/JNK pathway and apoptosis. Importantly, gadd45αa MO and p38/JNK inhibitors could partially rescue the defect of prmt6 morphants, the downstream targets of Prmt6, and the apoptosis ratios of the prmt6 morphants. Moreover, the results of ChIP quantitative real time PCR and luciferase reporter assay indicated that gadd45αa is a repressive target of Prmt6. Taken together, these results suggest that maternal Prmt6 is essential to early zebrafish development by directly repressing gadd45αa.
Keywords: apoptosis; c-Jun N-terminal kinase (JNK); gadd45αa; histone methylation; p38; prmt6; zebrafish.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
Similar articles
-
Genomic Location of PRMT6-Dependent H3R2 Methylation Is Linked to the Transcriptional Outcome of Associated Genes.Cell Rep. 2018 Sep 18;24(12):3339-3352. doi: 10.1016/j.celrep.2018.08.052. Cell Rep. 2018. PMID: 30232013
-
PRMT6 mediates CSE induced inflammation and apoptosis.Int Immunopharmacol. 2015 Jan;24(1):95-101. doi: 10.1016/j.intimp.2014.10.029. Epub 2014 Dec 4. Int Immunopharmacol. 2015. PMID: 25481537
-
The Arginine Methyltransferase PRMT6 Cooperates with Polycomb Proteins in Regulating HOXA Gene Expression.PLoS One. 2016 Feb 5;11(2):e0148892. doi: 10.1371/journal.pone.0148892. eCollection 2016. PLoS One. 2016. PMID: 26848759 Free PMC article.
-
Contributions of the histone arginine methyltransferase PRMT6 to the epigenetic function of RUNX1.Crit Rev Eukaryot Gene Expr. 2013;23(3):265-74. doi: 10.1615/critreveukaryotgeneexpr.2013007527. Crit Rev Eukaryot Gene Expr. 2013. PMID: 23879542 Review.
-
Structure, Activity and Function of the Protein Arginine Methyltransferase 6.Life (Basel). 2021 Sep 11;11(9):951. doi: 10.3390/life11090951. Life (Basel). 2021. PMID: 34575100 Free PMC article. Review.
Cited by
-
Histones, Their Variants and Post-translational Modifications in Zebrafish Development.Front Cell Dev Biol. 2020 Jun 5;8:456. doi: 10.3389/fcell.2020.00456. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 32582716 Free PMC article. Review.
-
Genome-Wide Characterization of Selection Signatures and Runs of Homozygosity in Ugandan Goat Breeds.Front Genet. 2018 Aug 14;9:318. doi: 10.3389/fgene.2018.00318. eCollection 2018. Front Genet. 2018. PMID: 30154830 Free PMC article.
-
The Emerging Role of PRMT6 in Cancer.Front Oncol. 2022 Mar 4;12:841381. doi: 10.3389/fonc.2022.841381. eCollection 2022. Front Oncol. 2022. PMID: 35311114 Free PMC article. Review.
-
Genetic evidence for partial redundancy between the arginine methyltransferases CARM1 and PRMT6.J Biol Chem. 2020 Dec 11;295(50):17060-17070. doi: 10.1074/jbc.RA120.014704. Epub 2020 Oct 2. J Biol Chem. 2020. PMID: 33008887 Free PMC article.
-
Protein arginine methyltransferases in protozoan parasites.Parasitology. 2022 Apr;149(4):427-435. doi: 10.1017/S0031182021002043. Epub 2021 Dec 6. Parasitology. 2022. PMID: 35331350 Free PMC article. Review.
References
-
- Kane D. A., and Kimmel C. B. (1993) The zebrafish midblastula transition. Development 119, 447–456 - PubMed
-
- Schier A. F. (2007) The maternal-zygotic transition: death and birth of RNAs. Science 316, 406–407 - PubMed
-
- Zhang W. W., Zhang Y. B., Zhao X. X., Hua Y., Wu Z. L., Yan Y. C., and Li Y. P. (2015) Prmt7 regulates epiboly by facilitating 2-OST and modulating actin cytoskeleton. J. Mol. Cell Biol. 7, 489–491 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
