

A Simple Method for Discovering Druggable, Specific Glycosaminoglycan-Protein Systems. Elucidation of Key Principles from Heparin/Heparan Sulfate-Binding Proteins
- PMID: 26488293
- PMCID: PMC4619353
- DOI: 10.1371/journal.pone.0141127
A Simple Method for Discovering Druggable, Specific Glycosaminoglycan-Protein Systems. Elucidation of Key Principles from Heparin/Heparan Sulfate-Binding Proteins
Abstract
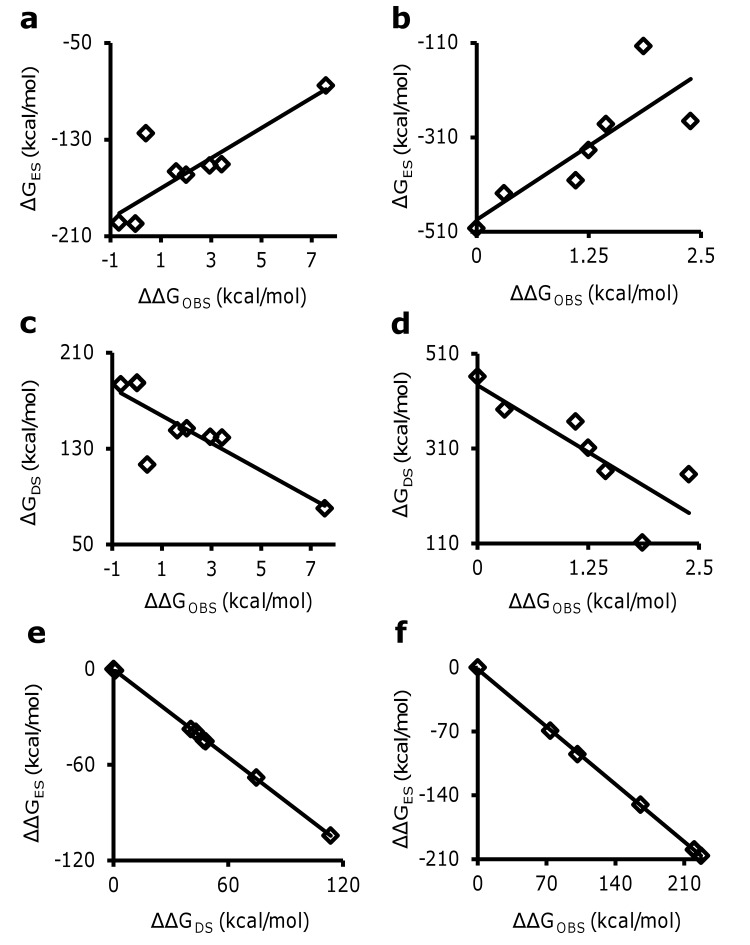
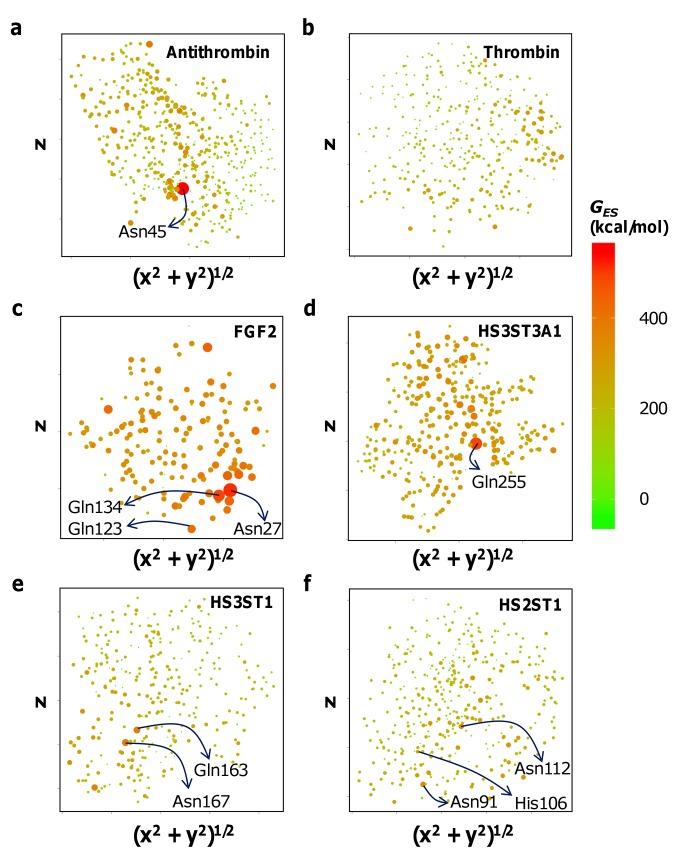
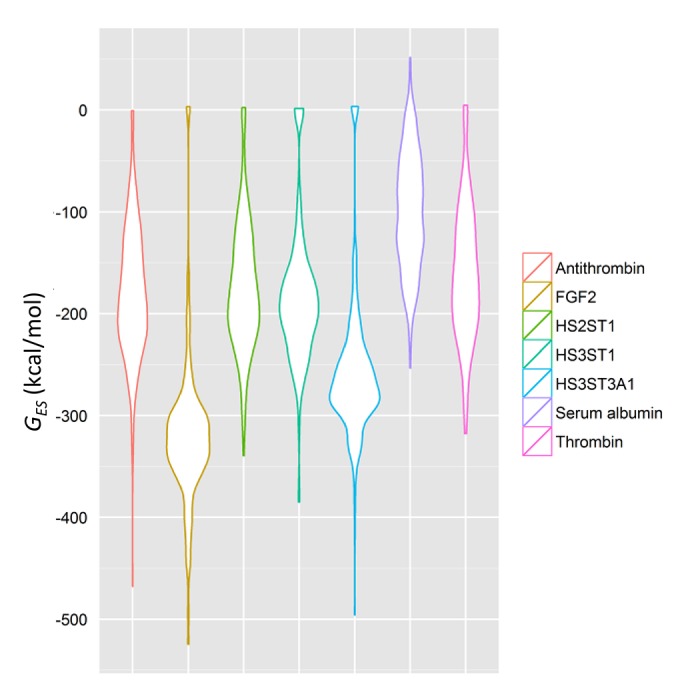
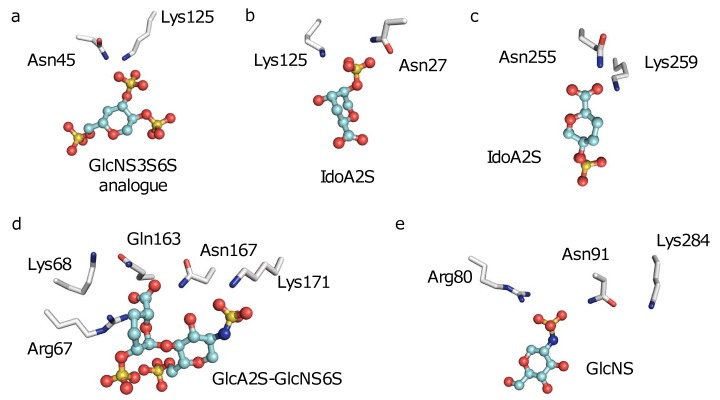
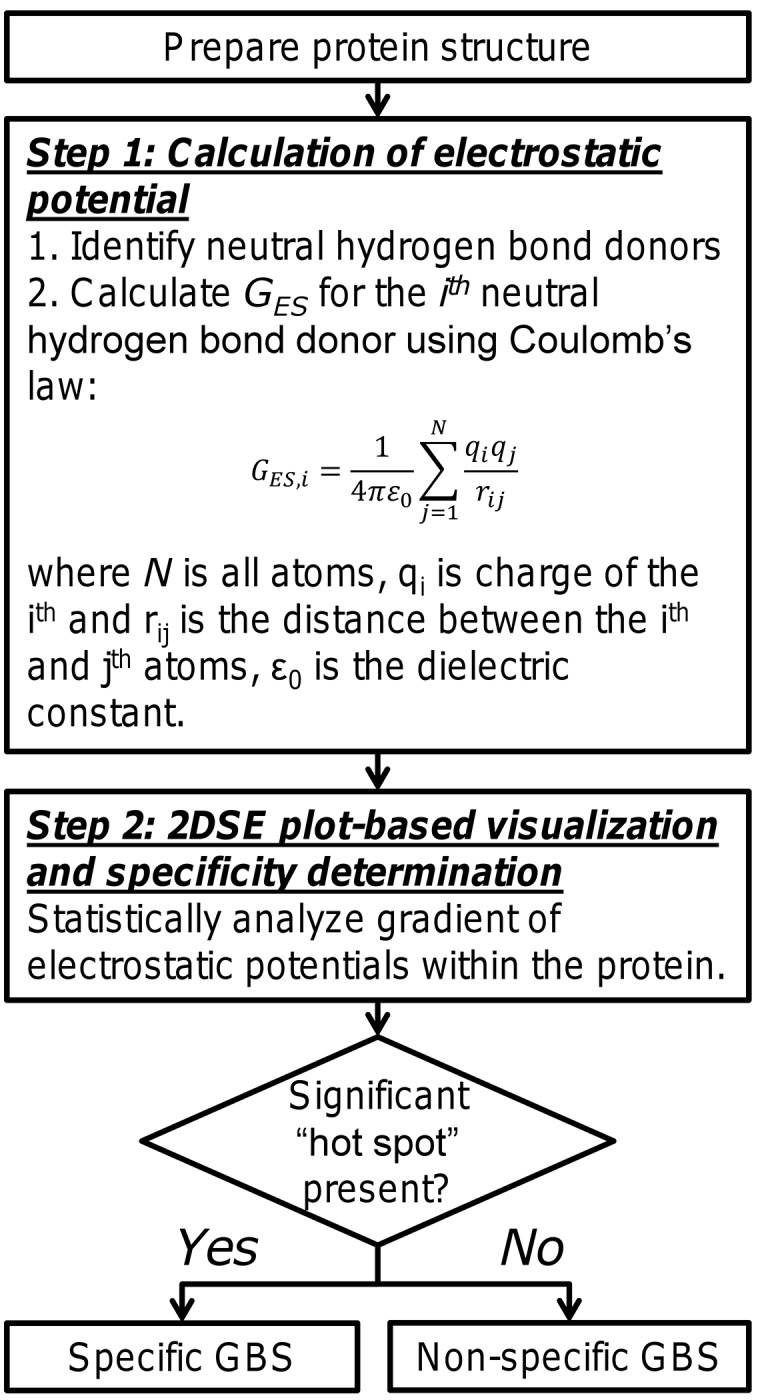
Glycosaminoglycans (GAGs) affect human physiology and pathology by modulating more than 500 proteins. GAG-protein interactions are generally assumed to be ionic and nonspecific, but specific interactions do exist. Here, we present a simple method to identify the GAG-binding site (GBS) on proteins that in turn helps predict high specific GAG-protein systems. Contrary to contemporary thinking, we found that the electrostatic potential at basic arginine and lysine residues neither identifies the GBS consistently, nor its specificity. GBSs are better identified by considering the potential at neutral hydrogen bond donors such as asparagine or glutamine sidechains. Our studies also reveal that an unusual constellation of ionic and non-ionic residues in the binding site leads to specificity. Nature engineers the local environment of Asn45 of antithrombin, Gln255 of 3-O-sulfotransferase 3, Gln163 and Asn167 of 3-O-sulfotransferase 1 and Asn27 of basic fibroblast growth factor in the respective GBSs to induce specificity. Such residues are distinct from other uncharged residues on the same protein structure in possessing a significantly higher electrostatic potential, resultant from the local topology. In contrast, uncharged residues on nonspecific GBSs such as thrombin and serum albumin possess a diffuse spread of electrostatic potential. Our findings also contradict the paradigm that GAG-binding sites are simply a collection of contiguous Arg/Lys residues. Our work demonstrates the basis for discovering specifically interacting and druggable GAG-protein systems based on the structure of protein alone, without requiring access to any structure-function relationship data.
Conflict of interest statement
Figures

References
-
- Sobel M, Soler DF, Kermode JC, Harris RB. Localization and characterization of a heparin binding domain peptide of human von Willebrand factor. J Biol Chem. 1992;267: 8857–62. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
