

ABCG1 regulates mouse adipose tissue macrophage cholesterol levels and ratio of M1 to M2 cells in obesity and caloric restriction
- PMID: 26489644
- PMCID: PMC4655989
- DOI: 10.1194/jlr.M063354
ABCG1 regulates mouse adipose tissue macrophage cholesterol levels and ratio of M1 to M2 cells in obesity and caloric restriction
Abstract
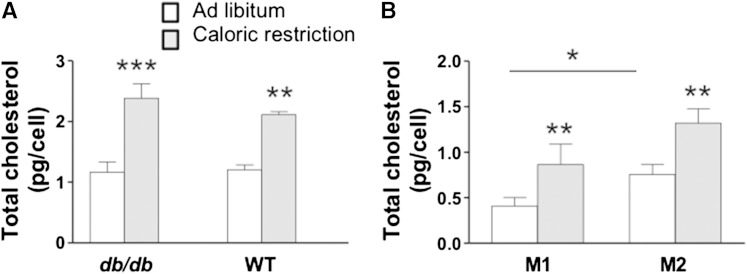
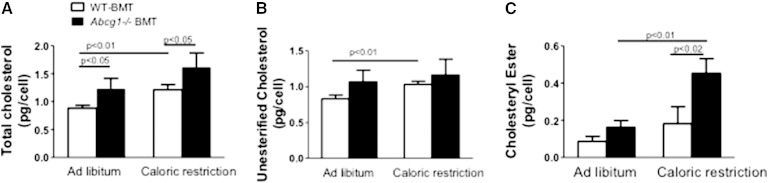
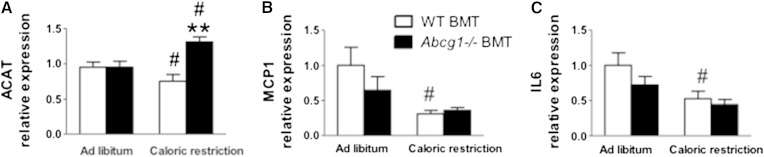
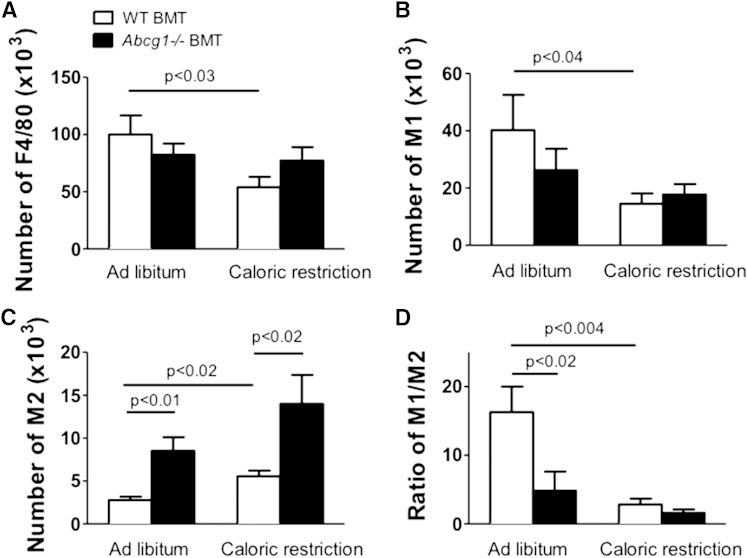
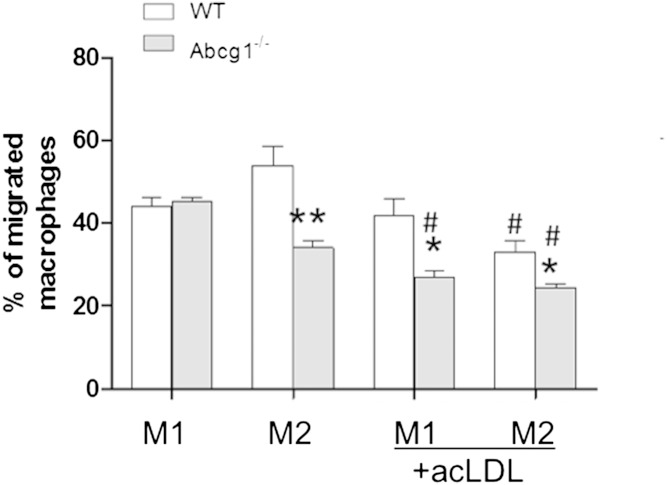
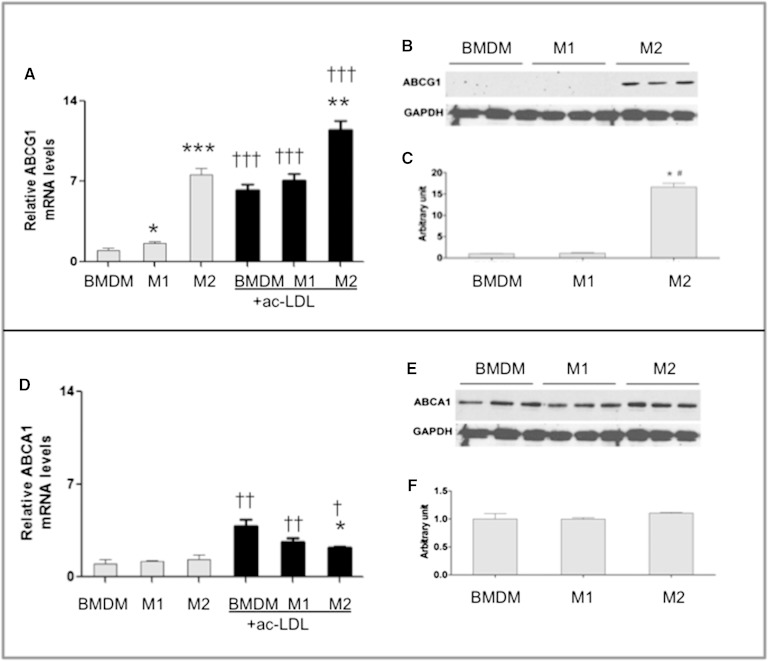
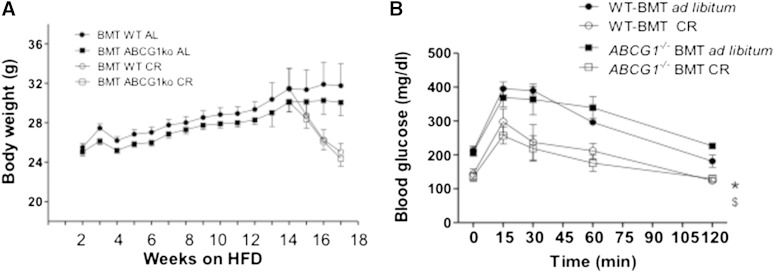
In addition to triacylglycerols, adipocytes contain a large reserve of unesterified cholesterol. During adipocyte lipolysis and cell death seen during severe obesity and weight loss, free fatty acids and cholesterol become available for uptake and processing by adipose tissue macrophages (ATMs). We hypothesize that ATMs become cholesterol enriched and participate in cholesterol clearance from adipose tissue. We previously showed that ABCG1 is robustly upregulated in ATMs taken from obese mice and further enhanced by caloric restriction. Here, we found that ATMs taken from obese and calorie-restricted mice derived from transplantation of WT or Abcg1-deficient bone marrow are cholesterol enriched. ABCG1 levels regulate the ratio of classically activated (M1) to alternatively activated (M2) ATMs and their cellular cholesterol content. Using WT and Abcg1(-/-) cultured macrophages, we found that Abcg1 is most highly expressed by M2 macrophages and that ABCG1 deficiency is sufficient to retard macrophage chemotaxis. However, changes in myeloid expression of Abcg1 did not protect mice from obesity or impaired glucose homeostasis. Overall, ABCG1 modulates ATM cholesterol content in obesity and weight loss regimes leading to an alteration in M1 to M2 ratio that we suggest is due to the extent of macrophage egress from adipose tissue.
Keywords: ATP binding cassette transporter A1; ATP binding cassette transporter G1; acyl-CoA:cholesterol acyltransferase; diabetes; fatty acid; gallstones; lipids; mice; nutrition; sterols.
Copyright © 2015 by the American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Azagury D. E., and Lautz D. B.. 2011. Obesity overview: epidemiology, health and financial impact, and guidelines for qualification for surgical therapy. Gastrointest. Endosc. Clin. N. Am. 21: 189–201. - PubMed
-
- Banim P. J., Luben R. N., Bulluck H., Sharp S. J., Wareham N. J., Khaw K. T., and Hart A. R.. 2011. The aetiology of symptomatic gallstones quantification of the effects of obesity, alcohol and serum lipids on risk. Epidemiological and biomarker data from a UK prospective cohort study (EPIC-Norfolk). Eur. J. Gastroenterol. Hepatol. 23: 733–740. - PubMed
-
- Malik V. S., Willett W. C., and Hu F. B.. 2013. Global obesity: trends, risk factors and policy implications. Nat. Rev. Endocrinol. 9: 13–27. - PubMed
-
- World Health Organization. 2009. Global Health Risks: Mortality and Burden of Disease Attributable to Selected Major Risks. World Health Organization, Geneva.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
