

Binding of alphaherpesvirus glycoprotein H to surface α4β1-integrins activates calcium-signaling pathways and induces phosphatidylserine exposure on the plasma membrane
- PMID: 26489864
- PMCID: PMC4620472
- DOI: 10.1128/mBio.01552-15
Binding of alphaherpesvirus glycoprotein H to surface α4β1-integrins activates calcium-signaling pathways and induces phosphatidylserine exposure on the plasma membrane
Abstract
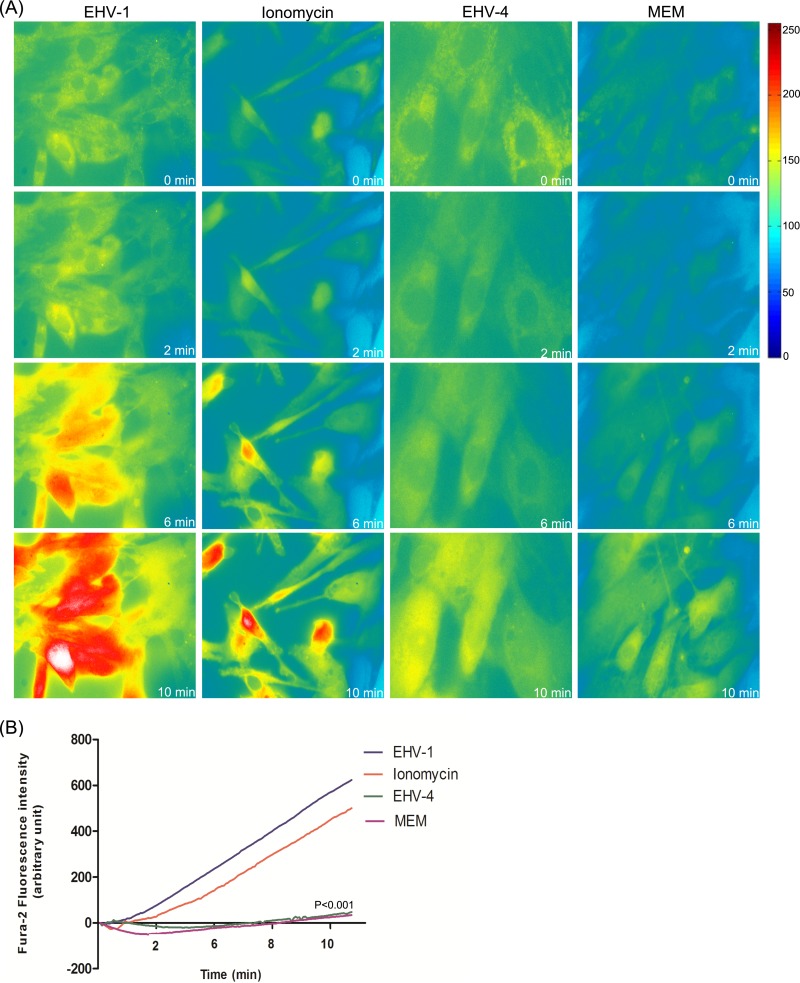
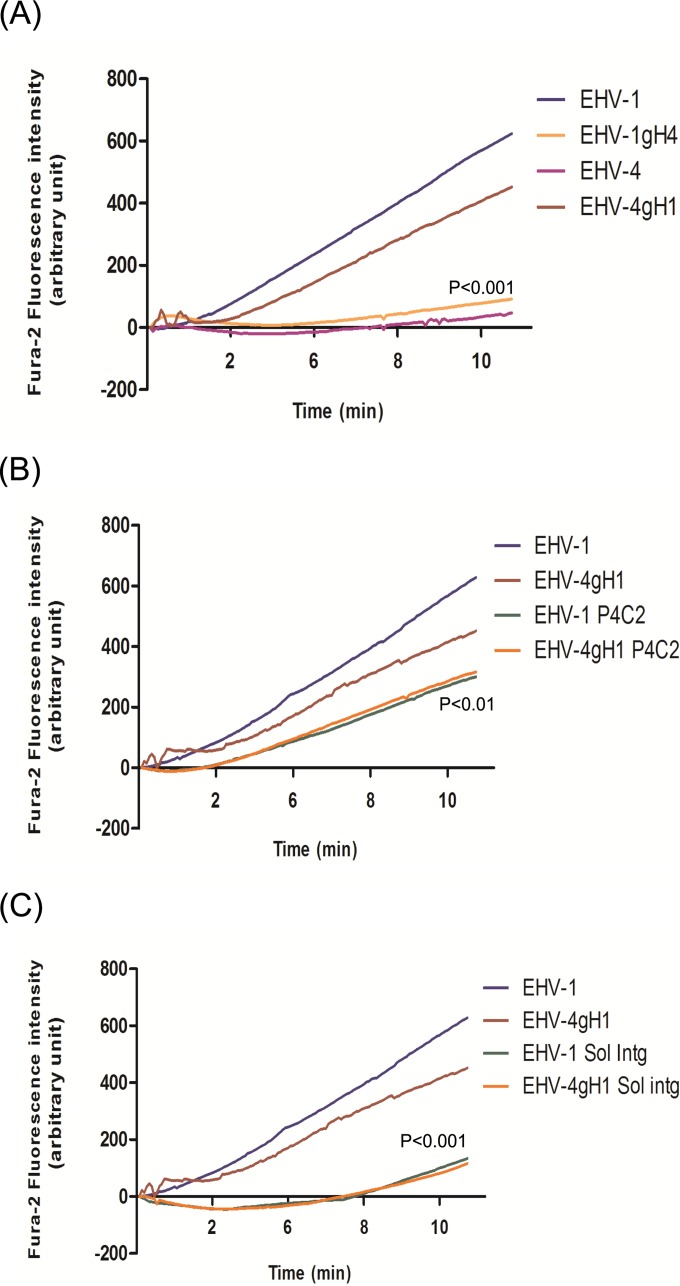
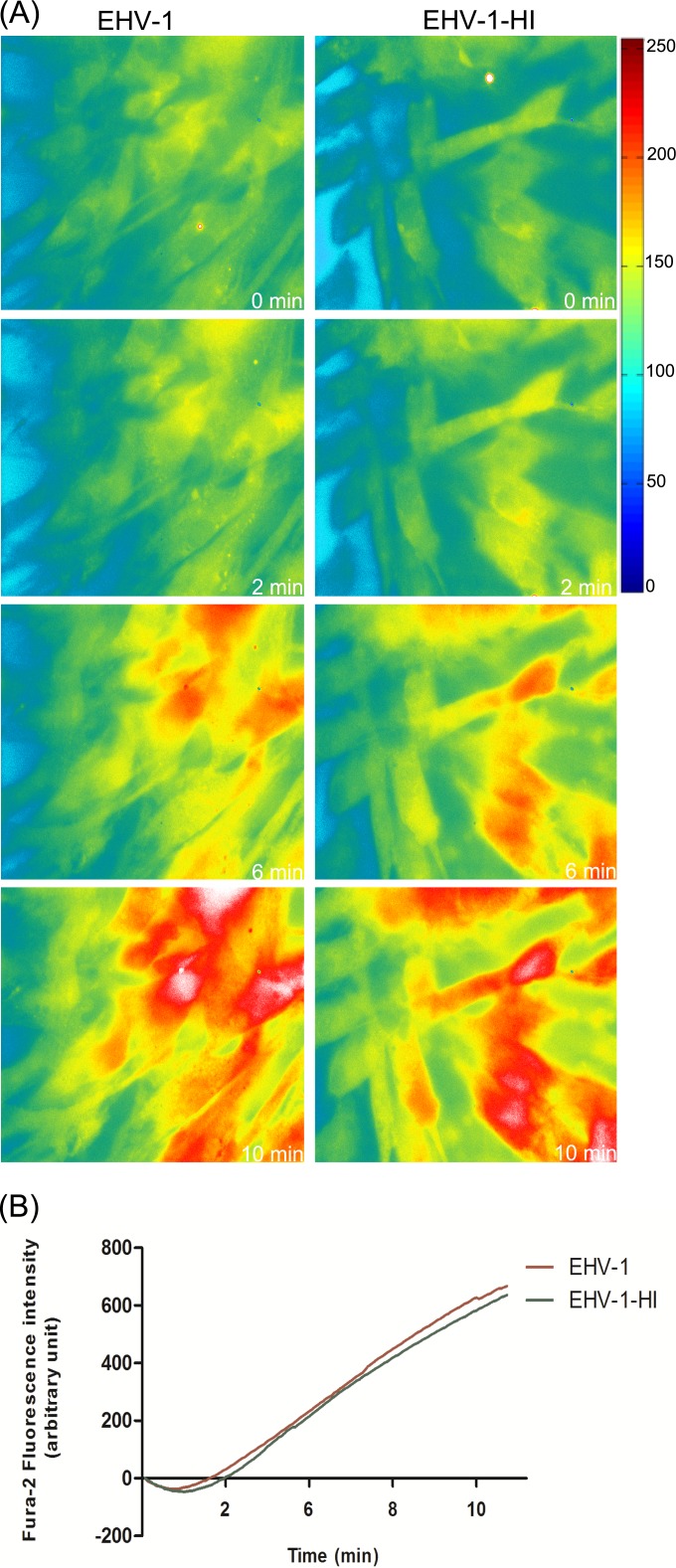
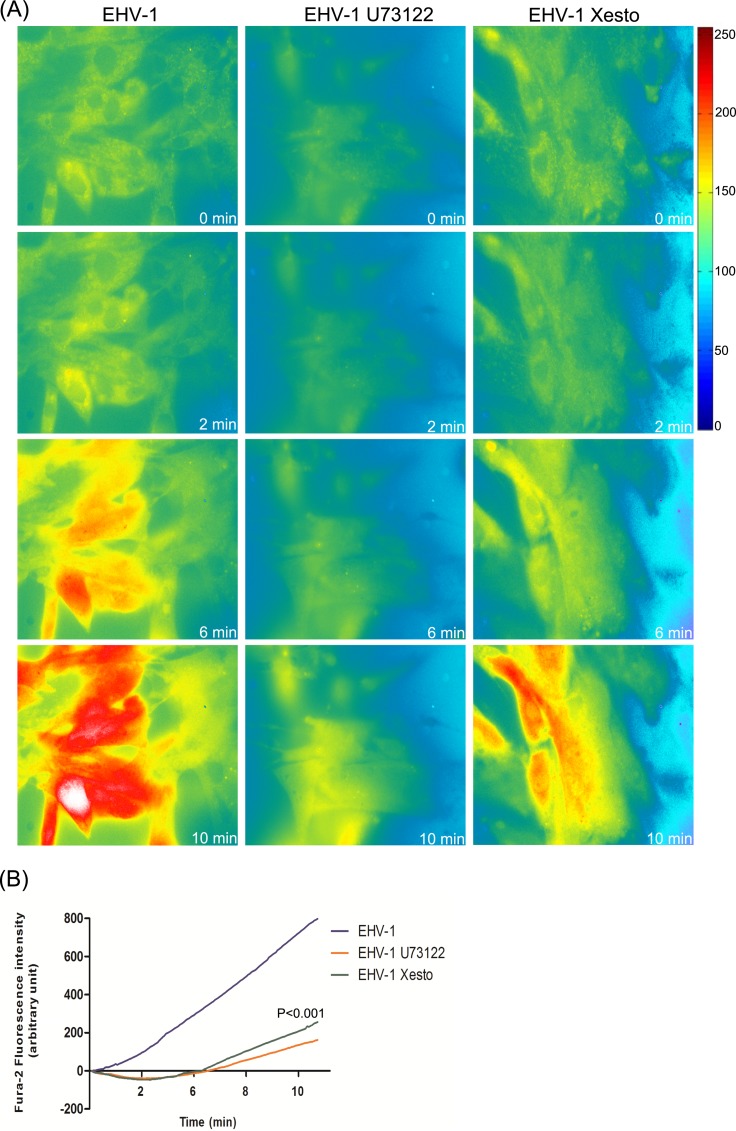
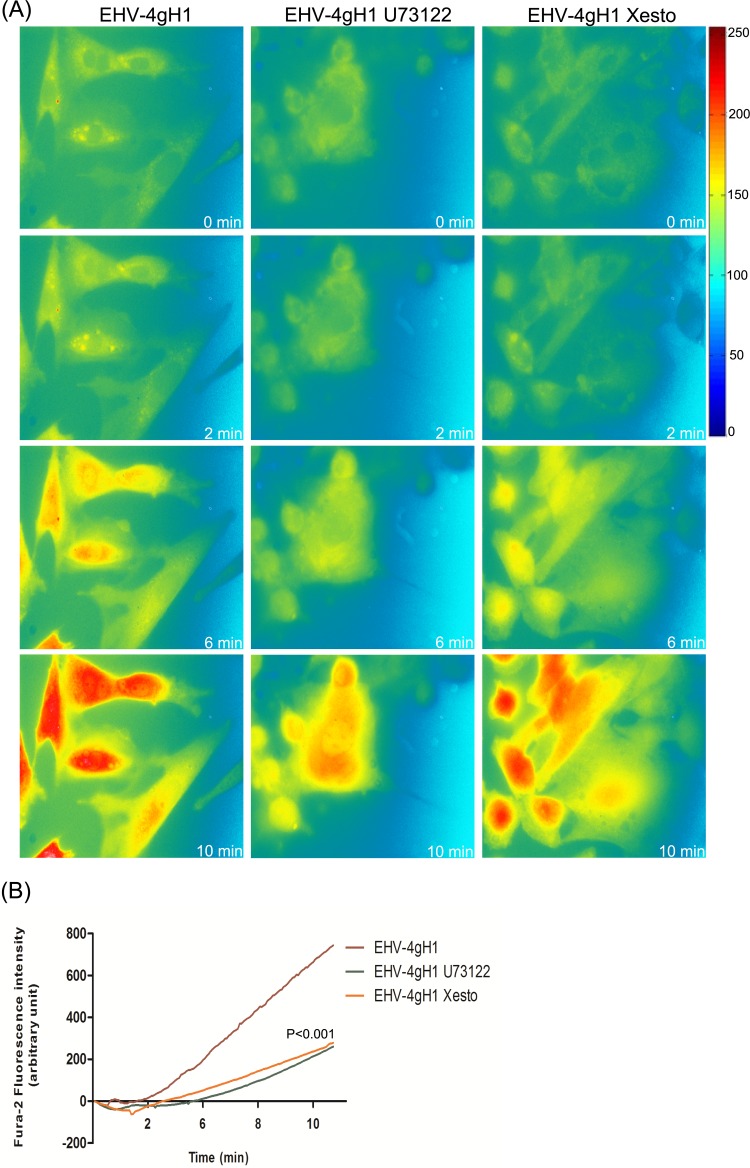
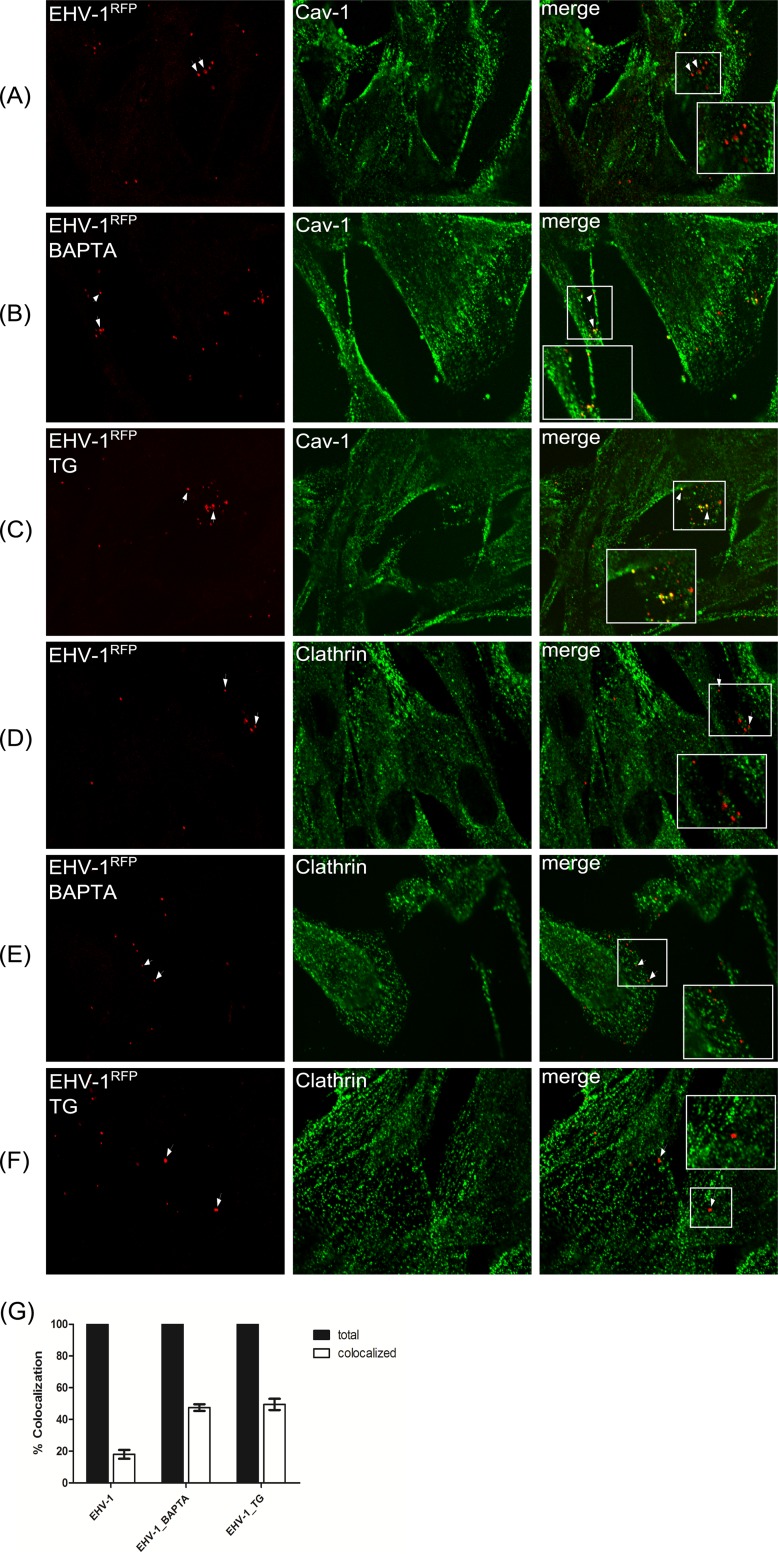
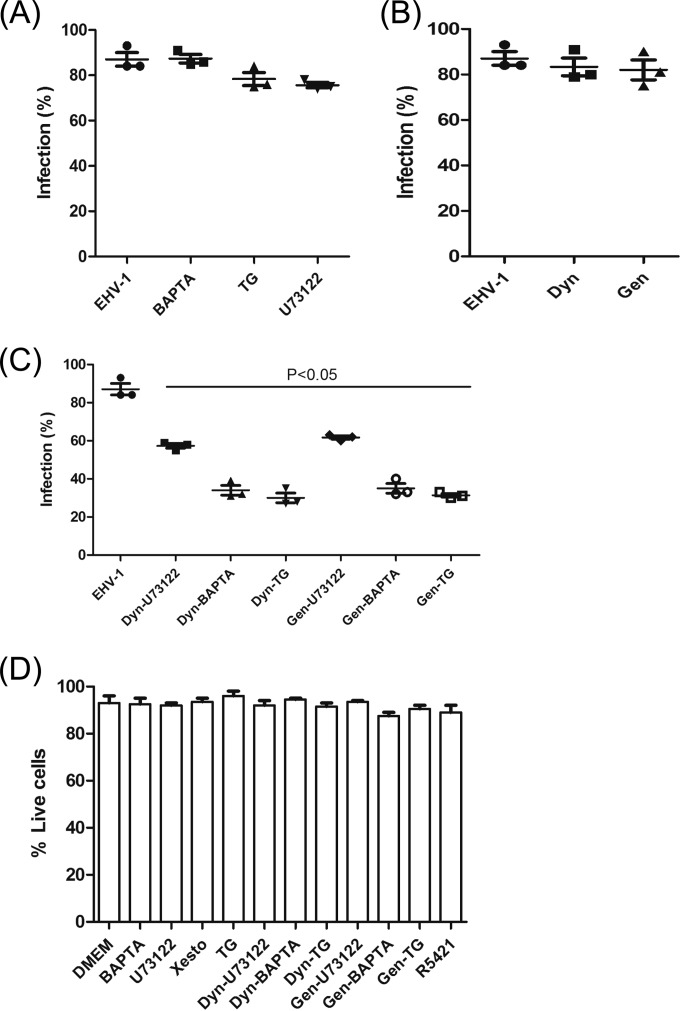
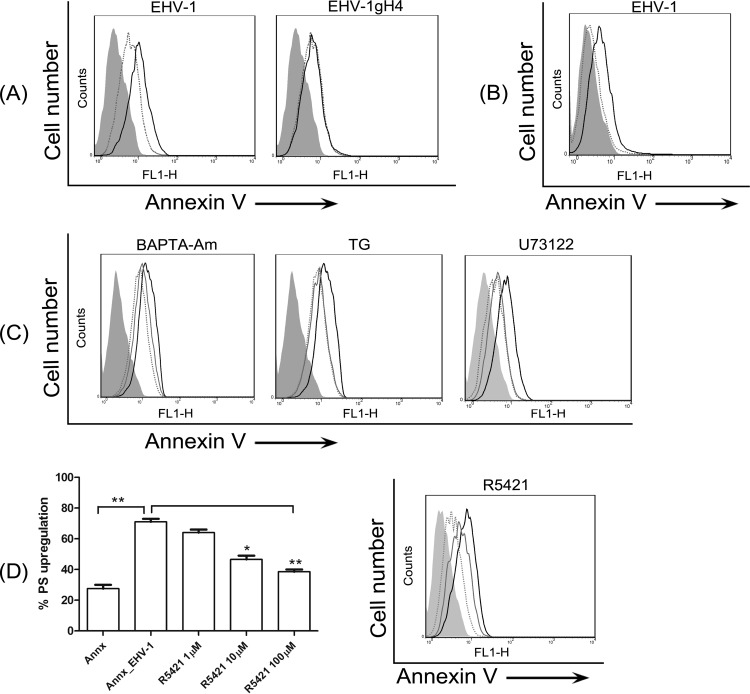
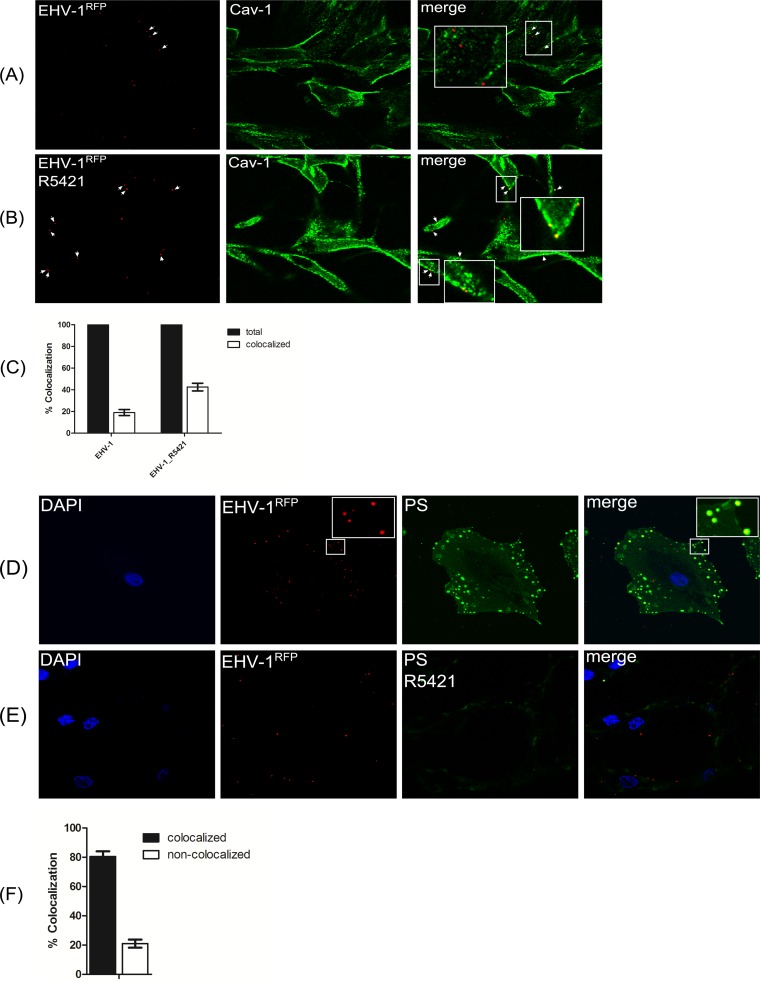
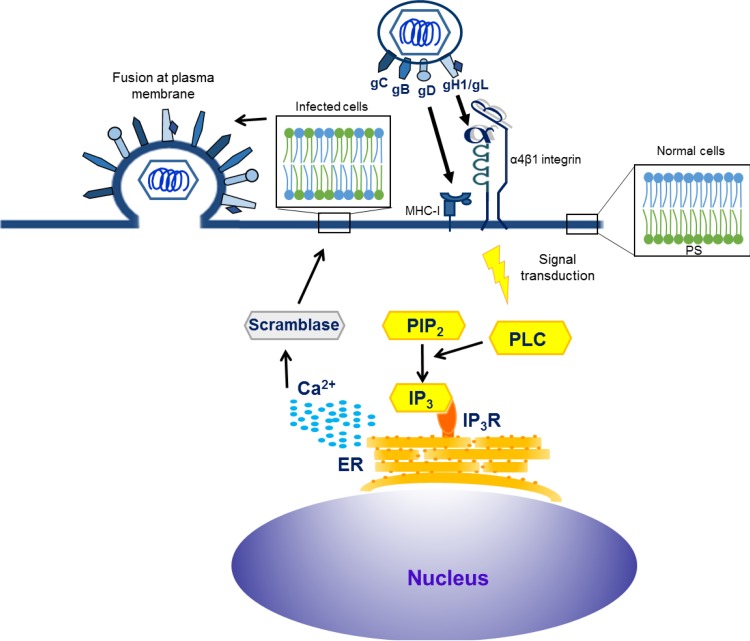
Intracellular signaling connected to integrin activation is known to induce cytoplasmic Ca(2+) release, which in turn mediates a number of downstream signals. The cellular entry pathways of two closely related alphaherpesviruses, equine herpesviruses 1 and 4 (EHV-1 and EHV-4), are differentially regulated with respect to the requirement of interaction of glycoprotein H (gH) with α4β1-integrins. We show here that binding of EHV-1, but not EHV-4, to target cells resulted in a rapid and significant increase in cytosolic Ca(2+) levels. EHV-1 expressing EHV-4 gH (gH4) in lieu of authentic gH1 failed to induce Ca(2+) release, while EHV-4 with gH1 triggered significant Ca(2+) release. Blocking the interaction between gH1 and α4β1-integrins, inhibiting phospholipase C (PLC) activation, or blocking binding of inositol 1,4,5-triphosphate (IP3) to its receptor on the endoplasmic reticulum (ER) abrogated Ca(2+) release. Interestingly, phosphatidylserine (PS) was exposed on the plasma membrane in response to cytosolic calcium increase after EHV-1 binding through a scramblase-dependent mechanism. Inhibition of both Ca(2+) release from the ER and scramblase activation blocked PS scrambling and redirected virus entry to the endocytic pathway, indicating that PS may play a role in facilitating virus entry directly at the plasma membrane.
Importance: Herpesviruses are a large family of enveloped viruses that infect a wide range of hosts, causing a variety of diseases. These viruses have developed a number of strategies for successful entry into different cell types. We and others have shown that alphaherpesviruses, including EHV-1 and herpes simplex virus 1 (HSV-1), can route their entry pathway and do so by manipulation of cell signaling cascades to ensure viral genome delivery to nuclei. We show here that the interaction between EHV-1 gH and cellular α4β1-integrins is necessary to induce emptying of ER calcium stores, which induces phosphatidylserine exposure on the plasma membrane through a scramblase-dependent mechanism. This change in lipid asymmetry facilitates virus entry and might help fusion of the viral envelope at the plasma membrane. These findings will help to advance our understanding of herpesvirus entry mechanism and may facilitate the development of novel drugs that can be implemented for prevention of infection and disease.
Copyright © 2015 Azab et al.
Figures
References
-
- Sasaki M, Hasebe R, Makino Y, Suzuki T, Fukushi H, Okamoto M, Matsuda K, Taniyama H, Sawa H, Kimura T. 2011. Equine major histocompatibility complex class I molecules act as entry receptors that bind to equine herpesvirus-1 glycoprotein D. Genes Cells 16:343–357. doi: 10.1111/j.1365-2443.2011.01491.x. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
