

Predator Stress-Induced CRF Release Causes Enduring Sensitization of Basolateral Amygdala Norepinephrine Systems that Promote PTSD-Like Startle Abnormalities
- PMID: 26490866
- PMCID: PMC4683687
- DOI: 10.1523/JNEUROSCI.5080-14.2015
Predator Stress-Induced CRF Release Causes Enduring Sensitization of Basolateral Amygdala Norepinephrine Systems that Promote PTSD-Like Startle Abnormalities
Abstract
The neurobiology of post-traumatic stress disorder (PTSD) remains unclear. Intense stress promotes PTSD, which has been associated with exaggerated startle and deficient sensorimotor gating. Here, we examined the long-term sequelae of a rodent model of traumatic stress (repeated predator exposure) on amygdala systems that modulate startle and prepulse inhibition (PPI), an operational measure of sensorimotor gating. We show in rodents that repeated psychogenic stress (predator) induces long-lasting sensitization of basolateral amygdala (BLA) noradrenergic (NE) receptors (α1) via a corticotropin-releasing factor receptor 1 (CRF-R1)-dependent mechanism, and that these CRF1 and NE α1 receptors are highly colocalized on presumptive excitatory output projection neurons of the BLA. A profile identical to that seen with predator exposure was produced in nonstressed rats by intra-BLA infusions of CRF (200 ng/0.5 μl), but not by repeated NE infusions (20 μg/0.5 μl). Infusions into the adjacent central nucleus of amygdala had no effect. Importantly, the predator stress- or CRF-induced sensitization of BLA manifested as heightened startle and PPI deficits in response to subsequent subthreshold NE system challenges (with intra-BLA infusions of 0.3 μg/0.5 μl NE), up to 1 month after stress. This profile of effects closely resembles aspects of PTSD. Hence, we reveal a discrete neural pathway mediating the enhancement of NE system function seen in PTSD, and we offer a model for characterizing potential new treatments that may work by modulating this BLA circuitry.
Significance statement: The present findings reveal a novel and discrete neural substrate that could underlie certain core deficits (startle and prepulse inhibition) that are observed in post-traumatic stress disorder (PTSD). It is shown here that repeated exposure to a rodent model of traumatic stress (predator exposure) produces a long-lasting sensitization of basolateral amygdala noradrenergic substrates [via a corticotropin-releasing factor (CRF)-dependent mechanism] that regulate startle, which is exaggerated in PTSD. Moreover, it is demonstrated that the sensitized noradrenergic receptors colocalize with CRF1 receptors on output projection neurons of the basolateral amygdala. Hence, this stress-induced sensitization of noradrenergic receptors on basolateral nucleus efferents has wide-ranging implications for the numerous deleterious sequelae of trauma exposure that are seen in multiple psychiatric illnesses, including PTSD.
Keywords: corticotropin-releasing factor; corticotropin-releasing hormone; noradrenergic; prepulse inhibition; schizophrenia; sensorimotor gating.
Copyright © 2015 the authors 0270-6474/15/3514270-16$15.00/0.
Figures
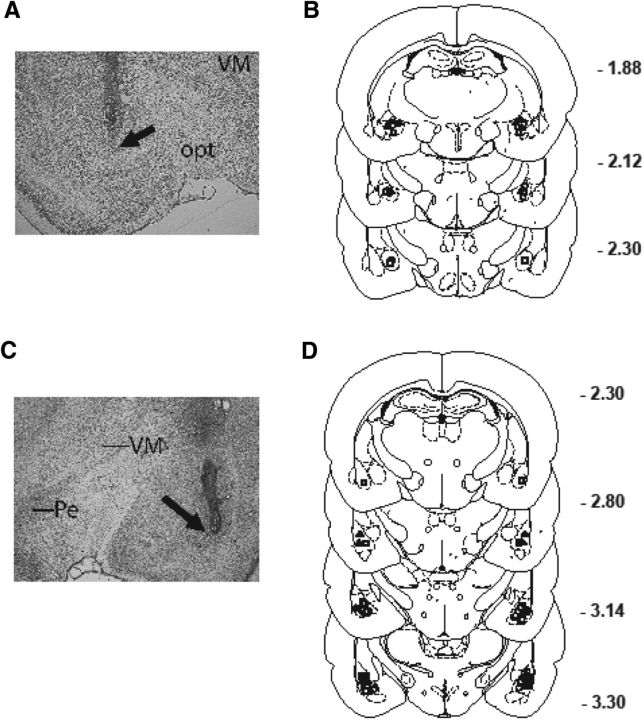
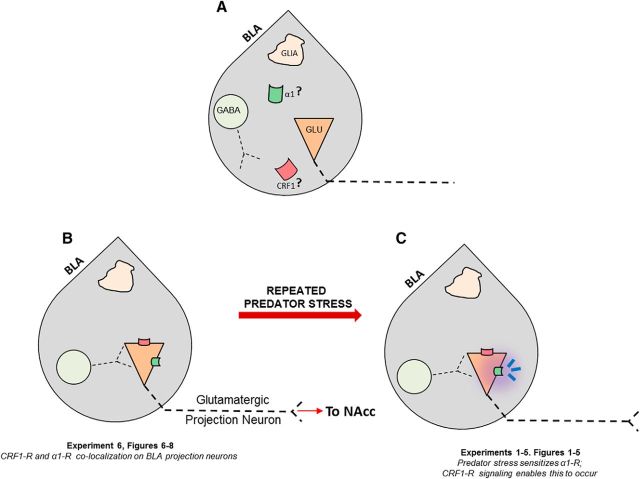
 ) and CRF1 (
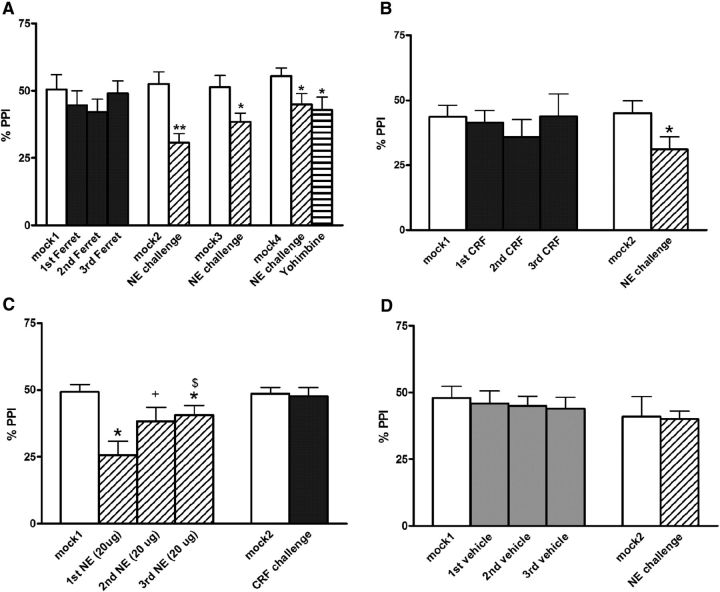
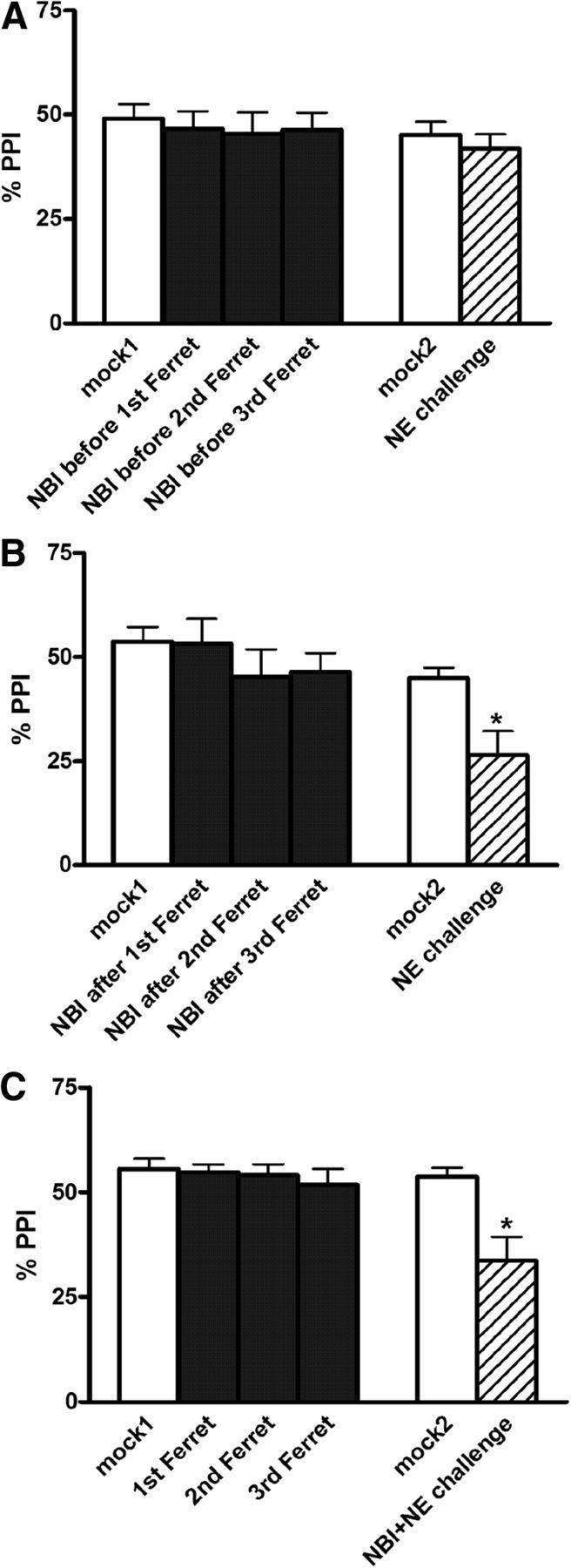
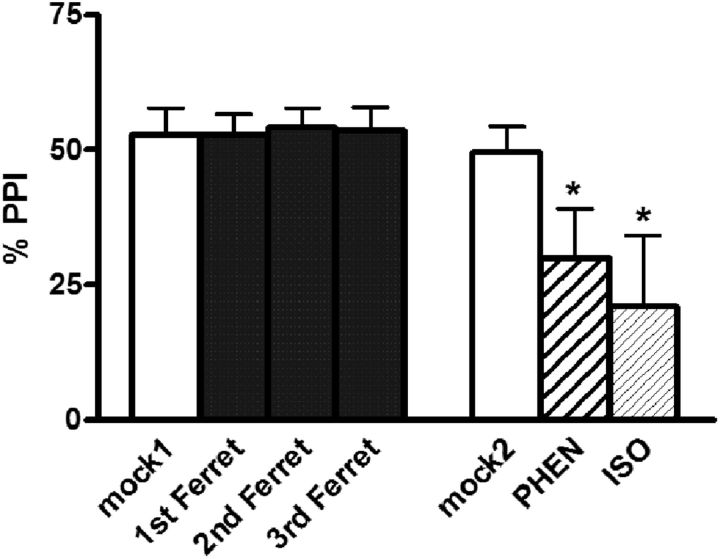
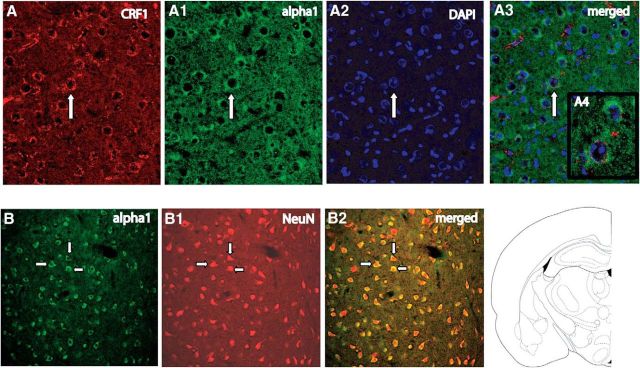
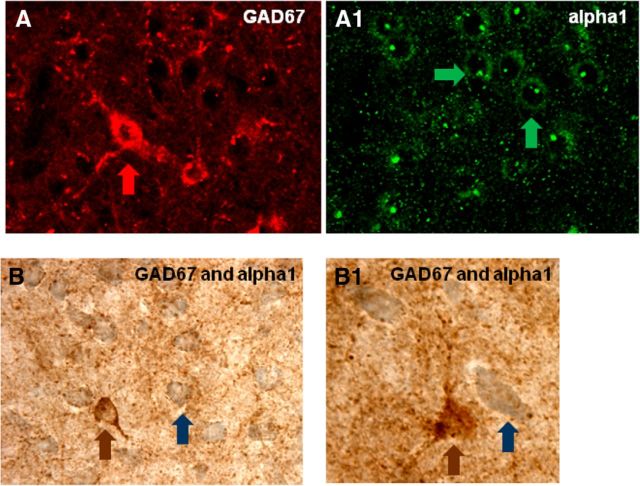
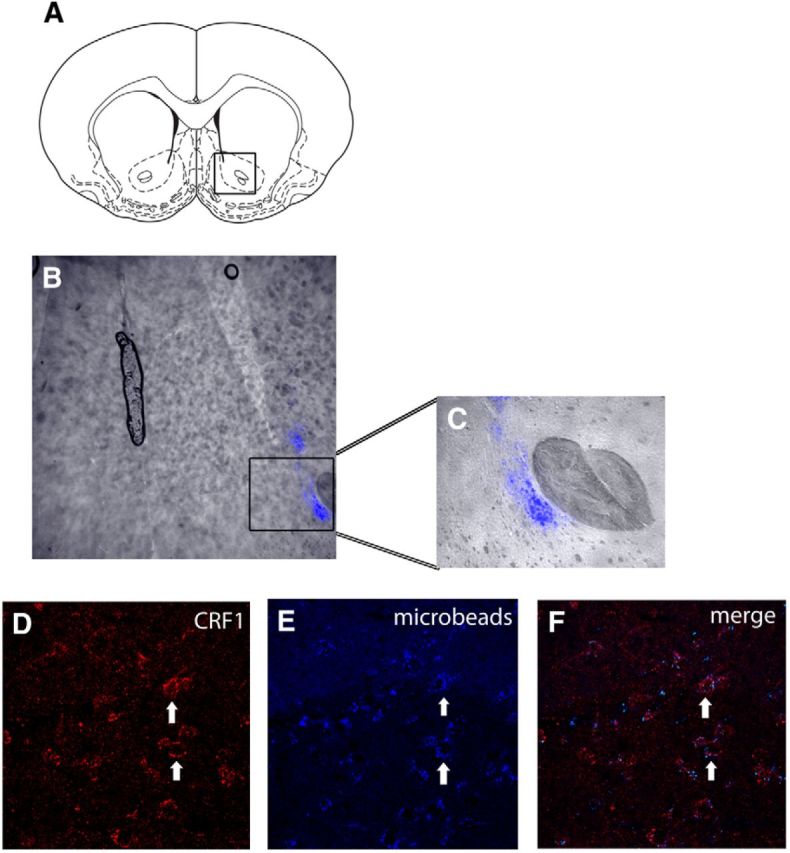
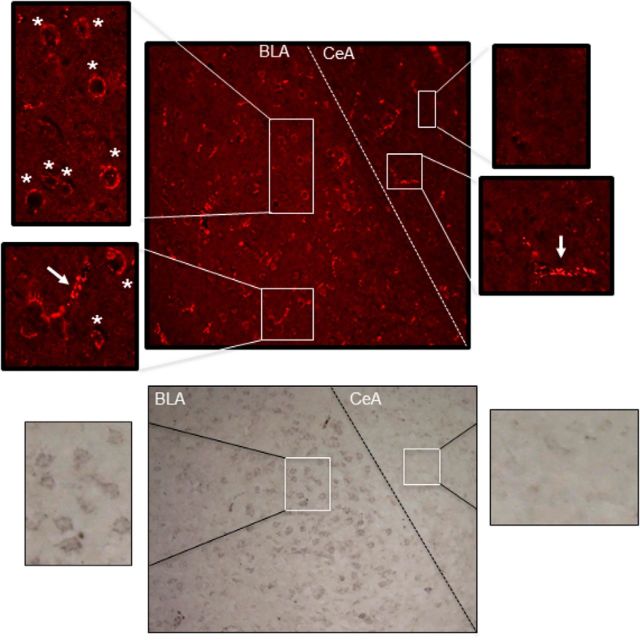
) and CRF1 ( ) receptors were both present in BLA, but their localization relative to each other and on different types of BLA cells (glia, orange; GABAergic inhibitory interneurons, blue; excitatory output neurons, green) was not known. B, We completed immunohistochemical and tract-tracing studies (Experiment 6; Figs. 6, 7, 8) to address these gaps in knowledge. First, we found that α1 and CRF1 receptors are colocalized on the same neurons and not on glial cells. Second, we failed to find these receptors on GABAergic cells. Third, we found that these receptors are present on BLA glutamatergic projection neurons that innervate the NAcc. Finally, we found that these output projection neurons that express CRF1 and α1 receptors are innervated by GABAergic neurons (presumptive inhibitory interneurons). C, Working model for behavioral sensitization induced by repeated predator exposure. Repeated exposure to predator stress sensitizes α1 receptors on BLA output neurons via initial activation of CRF1 receptors. Sensitized α1 receptors are hence abnormally responsive to low levels of NE, leading to increased glutamate release in downstream targets such as the NAcc, and thereby disrupting PPI. This putative mechanism of stress-induced BLA NE α1 receptor sensitization could thus contribute to startle abnormalities (exaggerated startle and disrupted PPI) that are seen in post-traumatic stress disorder.
) receptors were both present in BLA, but their localization relative to each other and on different types of BLA cells (glia, orange; GABAergic inhibitory interneurons, blue; excitatory output neurons, green) was not known. B, We completed immunohistochemical and tract-tracing studies (Experiment 6; Figs. 6, 7, 8) to address these gaps in knowledge. First, we found that α1 and CRF1 receptors are colocalized on the same neurons and not on glial cells. Second, we failed to find these receptors on GABAergic cells. Third, we found that these receptors are present on BLA glutamatergic projection neurons that innervate the NAcc. Finally, we found that these output projection neurons that express CRF1 and α1 receptors are innervated by GABAergic neurons (presumptive inhibitory interneurons). C, Working model for behavioral sensitization induced by repeated predator exposure. Repeated exposure to predator stress sensitizes α1 receptors on BLA output neurons via initial activation of CRF1 receptors. Sensitized α1 receptors are hence abnormally responsive to low levels of NE, leading to increased glutamate release in downstream targets such as the NAcc, and thereby disrupting PPI. This putative mechanism of stress-induced BLA NE α1 receptor sensitization could thus contribute to startle abnormalities (exaggerated startle and disrupted PPI) that are seen in post-traumatic stress disorder.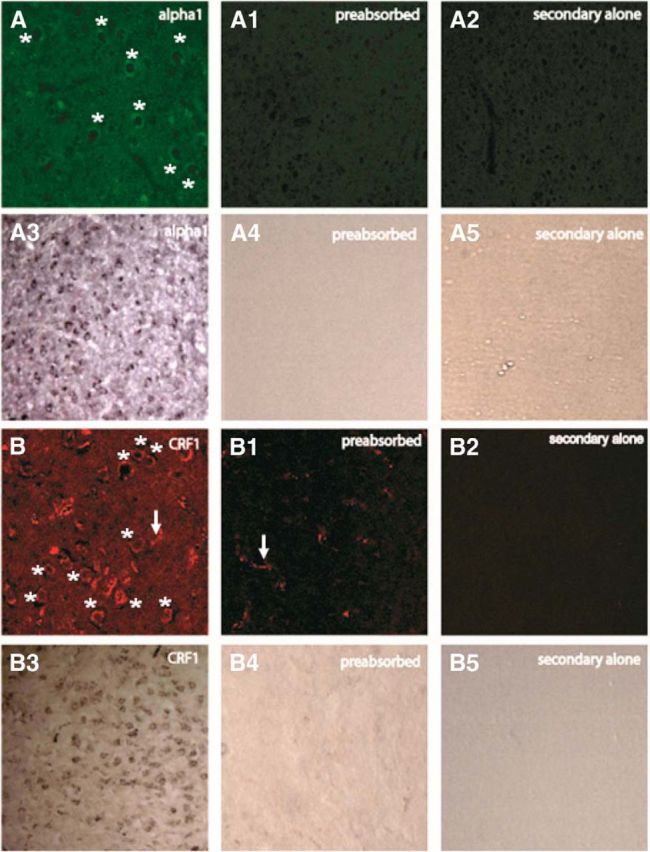
Similar articles
-
Repeated norepinephrine receptor stimulation in the BNST induces sensorimotor gating deficits via corticotropin releasing factor.Neuropharmacology. 2020 Aug 1;172:108090. doi: 10.1016/j.neuropharm.2020.108090. Epub 2020 Apr 29. Neuropharmacology. 2020. PMID: 32360378 Free PMC article.
-
Enduring sensorimotor gating abnormalities following predator exposure or corticotropin-releasing factor in rats: a model for PTSD-like information-processing deficits?Neuropharmacology. 2012 Feb;62(2):737-48. doi: 10.1016/j.neuropharm.2011.01.040. Epub 2011 Feb 1. Neuropharmacology. 2012. PMID: 21288473 Free PMC article.
-
Corticotropin-releasing factor and noradrenergic signalling exert reciprocal control over startle reactivity.Int J Neuropsychopharmacol. 2011 Oct;14(9):1179-94. doi: 10.1017/S1461145710001409. Epub 2010 Dec 21. Int J Neuropsychopharmacol. 2011. PMID: 21205416 Free PMC article.
-
The center of the emotional universe: Alcohol, stress, and CRF1 amygdala circuitry.Alcohol. 2018 Nov;72:61-73. doi: 10.1016/j.alcohol.2018.03.009. Epub 2018 Mar 30. Alcohol. 2018. PMID: 30220589 Free PMC article. Review.
-
The Locus Coeruleus: Anatomy, Physiology, and Stress-Related Neuropsychiatric Disorders.Eur J Neurosci. 2025 Apr;61(7):e70111. doi: 10.1111/ejn.70111. Eur J Neurosci. 2025. PMID: 40219735 Free PMC article. Review.
Cited by
-
Persistent Stress-Induced Neuroplastic Changes in the Locus Coeruleus/Norepinephrine System.Neural Plast. 2018 Jun 13;2018:1892570. doi: 10.1155/2018/1892570. eCollection 2018. Neural Plast. 2018. PMID: 30008741 Free PMC article. Review.
-
Stress-Induced Functional Alterations in Amygdala: Implications for Neuropsychiatric Diseases.Front Neurosci. 2018 May 29;12:367. doi: 10.3389/fnins.2018.00367. eCollection 2018. Front Neurosci. 2018. PMID: 29896088 Free PMC article. Review.
-
Ready for translation: non-invasive auricular vagus nerve stimulation inhibits psychophysiological indices of stimulus-specific fear and facilitates responding to repeated exposure in phobic individuals.Transl Psychiatry. 2025 Apr 9;15(1):135. doi: 10.1038/s41398-025-03352-0. Transl Psychiatry. 2025. PMID: 40204704 Free PMC article.
-
Locus Ceruleus Norepinephrine Release: A Central Regulator of CNS Spatio-Temporal Activation?Front Synaptic Neurosci. 2016 Aug 26;8:25. doi: 10.3389/fnsyn.2016.00025. eCollection 2016. Front Synaptic Neurosci. 2016. PMID: 27616990 Free PMC article.
-
Repeated norepinephrine receptor stimulation in the BNST induces sensorimotor gating deficits via corticotropin releasing factor.Neuropharmacology. 2020 Aug 1;172:108090. doi: 10.1016/j.neuropharm.2020.108090. Epub 2020 Apr 29. Neuropharmacology. 2020. PMID: 32360378 Free PMC article.
References
-
- Baker DG, West SA, Nicholson WE, Ekhator NN, Kasckow JW, Hill KK, Bruce AB, Orth DN, Geracioti TD., Jr Serial CSF corticotropin-releasing hormone levels and adrenocortical activity in combat veterans with posttraumatic stress disorder. Am J Psychiatry. 1999;156:585–588. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical