

Crystal Structures of a Piscine Betanodavirus: Mechanisms of Capsid Assembly and Viral Infection
- PMID: 26491970
- PMCID: PMC4619592
- DOI: 10.1371/journal.ppat.1005203
Crystal Structures of a Piscine Betanodavirus: Mechanisms of Capsid Assembly and Viral Infection
Abstract
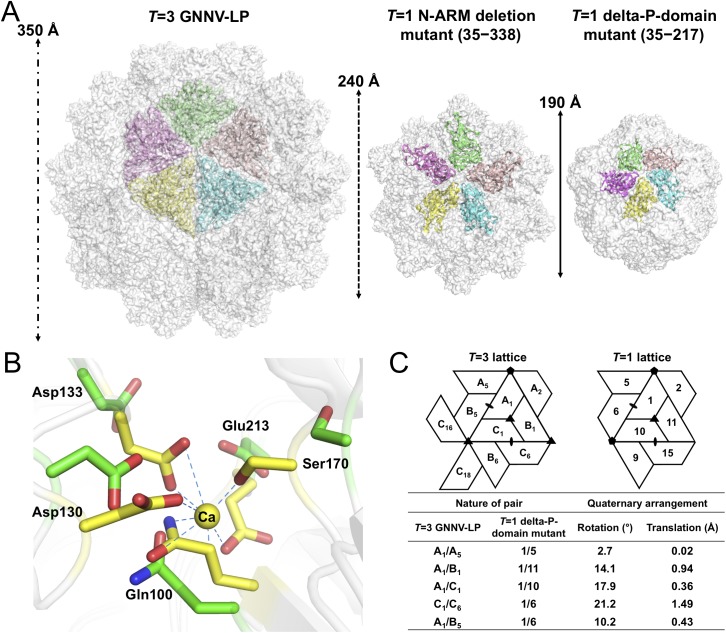
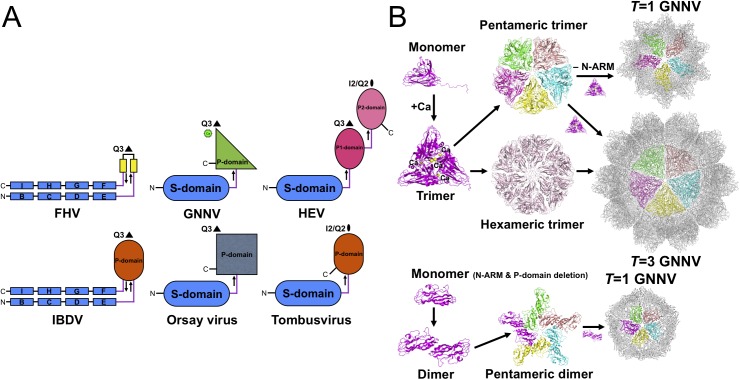
Betanodaviruses cause massive mortality in marine fish species with viral nervous necrosis. The structure of a T = 3 Grouper nervous necrosis virus-like particle (GNNV-LP) is determined by the ab initio method with non-crystallographic symmetry averaging at 3.6 Å resolution. Each capsid protein (CP) shows three major domains: (i) the N-terminal arm, an inter-subunit extension at the inner surface; (ii) the shell domain (S-domain), a jelly-roll structure; and (iii) the protrusion domain (P-domain) formed by three-fold trimeric protrusions. In addition, we have determined structures of the T = 1 subviral particles (SVPs) of (i) the delta-P-domain mutant (residues 35-217) at 3.1 Å resolution; and (ii) the N-ARM deletion mutant (residues 35-338) at 7 Å resolution; and (iii) the structure of the individual P-domain (residues 214-338) at 1.2 Å resolution. The P-domain reveals a novel DxD motif asymmetrically coordinating two Ca2+ ions, and seems to play a prominent role in the calcium-mediated trimerization of the GNNV CPs during the initial capsid assembly process. The flexible N-ARM (N-terminal arginine-rich motif) appears to serve as a molecular switch for T = 1 or T = 3 assembly. Finally, we find that polyethylene glycol, which is incorporated into the P-domain during the crystallization process, enhances GNNV infection. The present structural studies together with the biological assays enhance our understanding of the role of the P-domain of GNNV in the capsid assembly and viral infection by this betanodavirus.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



References
-
- Ahlquist P. RNA-dependent RNA polymerases, viruses, and RNA silencing. Science. 2002;296: 1270–1273. - PubMed
-
- Chao JA, Lee JH, Chapados BR, Debler EW, Schneemann A, Williamson JR. Dual modes of RNA-silencing suppression by Flock house virus protein B2. Nat Struct Mol Biol. 2006;12: 952–957. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
