

Modulation of Ambient Temperature-Dependent Flowering in Arabidopsis thaliana by Natural Variation of FLOWERING LOCUS M
- PMID: 26492483
- PMCID: PMC4619661
- DOI: 10.1371/journal.pgen.1005588
Modulation of Ambient Temperature-Dependent Flowering in Arabidopsis thaliana by Natural Variation of FLOWERING LOCUS M
Abstract
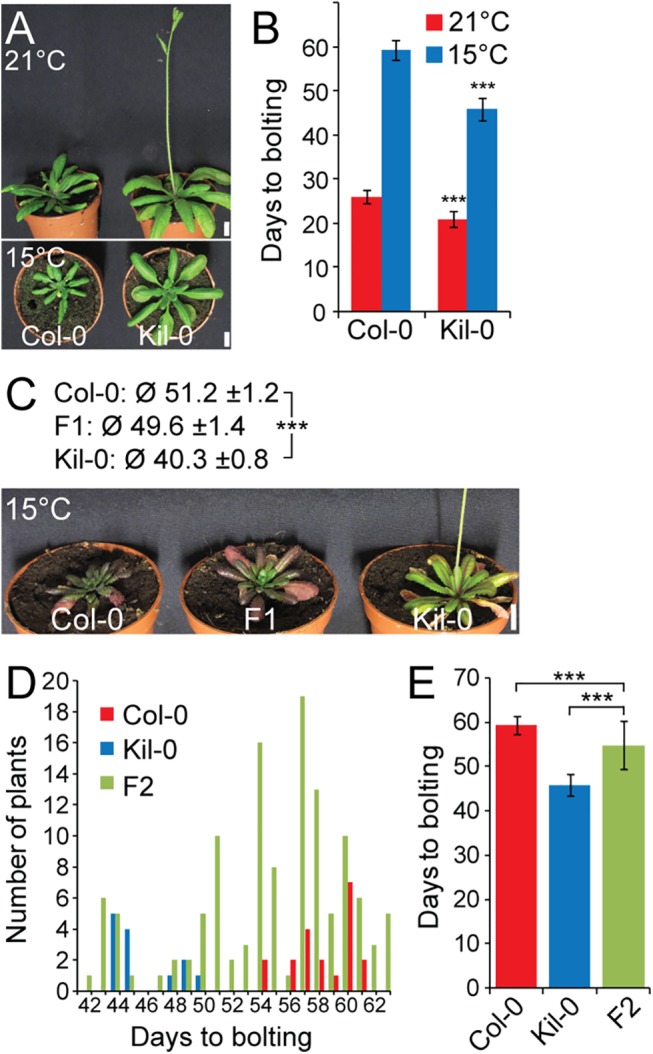
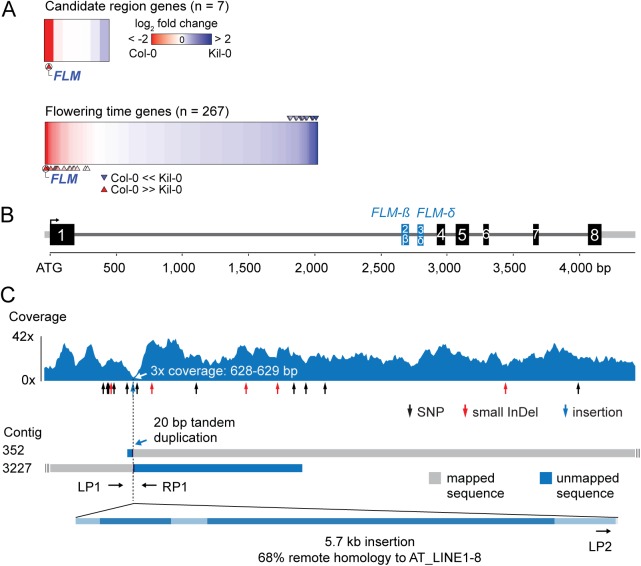
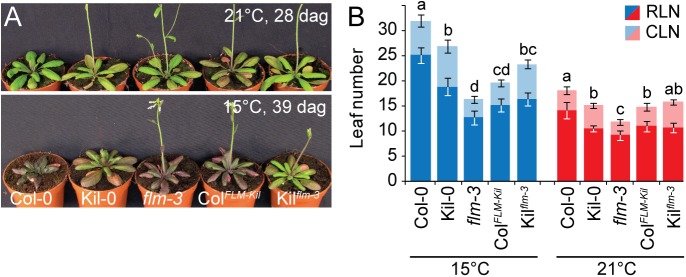
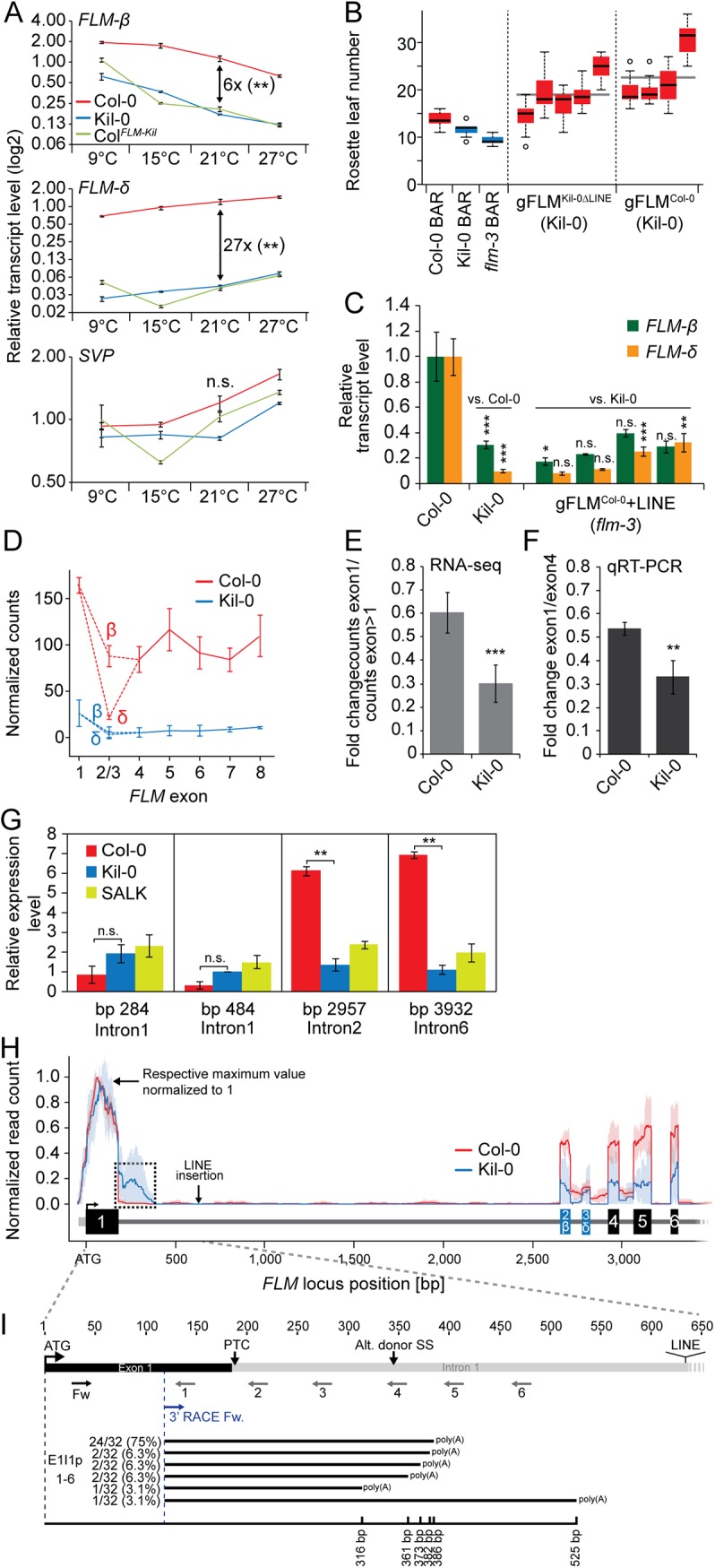
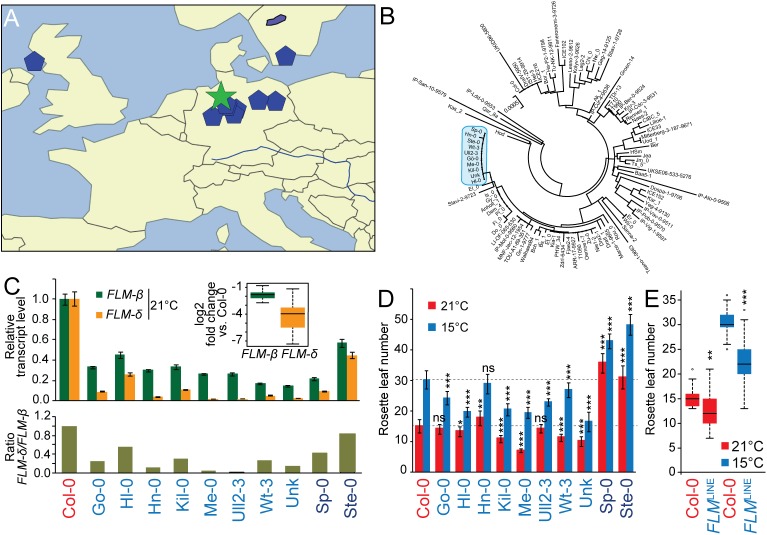
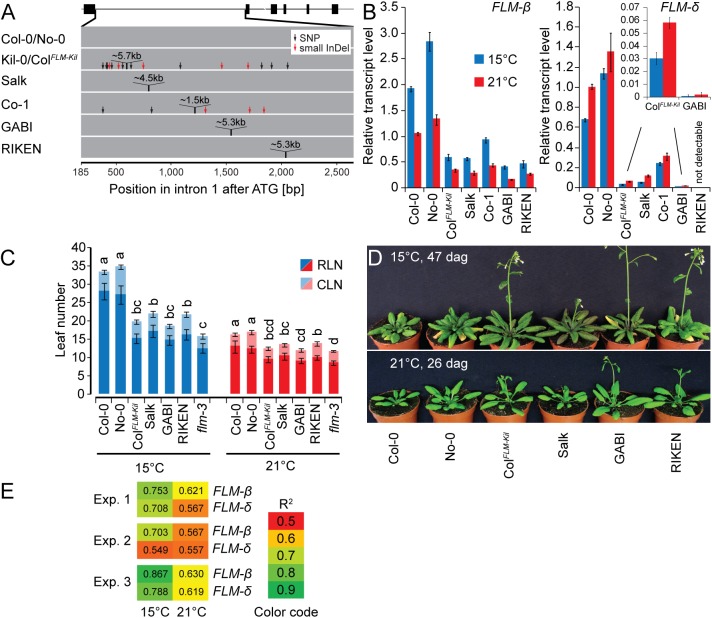
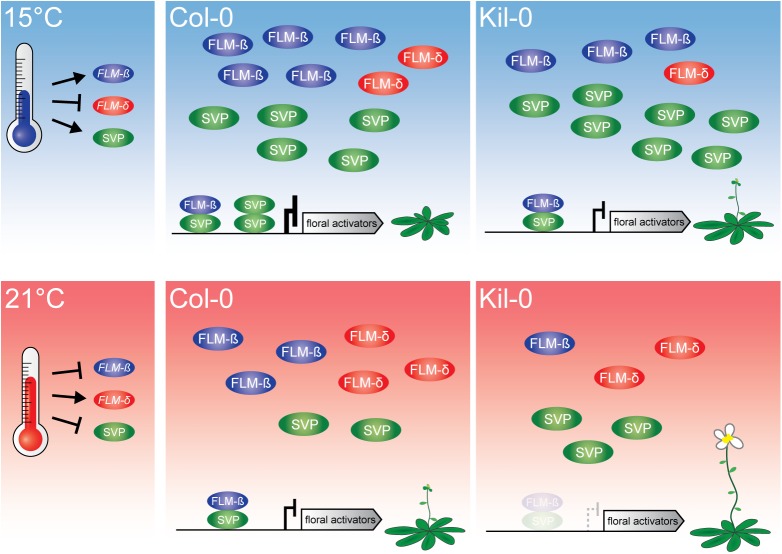
Plants integrate seasonal cues such as temperature and day length to optimally adjust their flowering time to the environment. Compared to the control of flowering before and after winter by the vernalization and day length pathways, mechanisms that delay or promote flowering during a transient cool or warm period, especially during spring, are less well understood. Due to global warming, understanding this ambient temperature pathway has gained increasing importance. In Arabidopsis thaliana, FLOWERING LOCUS M (FLM) is a critical flowering regulator of the ambient temperature pathway. FLM is alternatively spliced in a temperature-dependent manner and the two predominant splice variants, FLM-ß and FLM-δ, can repress and activate flowering in the genetic background of the A. thaliana reference accession Columbia-0. The relevance of this regulatory mechanism for the environmental adaptation across the entire range of the species is, however, unknown. Here, we identify insertion polymorphisms in the first intron of FLM as causative for accelerated flowering in many natural A. thaliana accessions, especially in cool (15°C) temperatures. We present evidence for a potential adaptive role of this structural variation and link it specifically to changes in the abundance of FLM-ß. Our results may allow predicting flowering in response to ambient temperatures in the Brassicaceae.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Comment in
-
Plasticity versus Adaptation of Ambient-Temperature Flowering Response.Trends Plant Sci. 2016 Jan;21(1):6-8. doi: 10.1016/j.tplants.2015.11.015. Epub 2015 Dec 14. Trends Plant Sci. 2016. PMID: 26698930
Similar articles
-
Natural haplotypes of FLM non-coding sequences fine-tune flowering time in ambient spring temperatures in Arabidopsis.Elife. 2017 Mar 15;6:e22114. doi: 10.7554/eLife.22114. Elife. 2017. PMID: 28294941 Free PMC article.
-
Contribution of major FLM isoforms to temperature-dependent flowering in Arabidopsis thaliana.J Exp Bot. 2017 Nov 2;68(18):5117-5127. doi: 10.1093/jxb/erx328. J Exp Bot. 2017. PMID: 29036339 Free PMC article.
-
MAF2 Is Regulated by Temperature-Dependent Splicing and Represses Flowering at Low Temperatures in Parallel with FLM.PLoS One. 2015 May 8;10(5):e0126516. doi: 10.1371/journal.pone.0126516. eCollection 2015. PLoS One. 2015. PMID: 25955034 Free PMC article.
-
Flowering Locus C's Lessons: Conserved Chromatin Switches Underpinning Developmental Timing and Adaptation.Plant Physiol. 2015 Aug;168(4):1237-45. doi: 10.1104/pp.15.00496. Epub 2015 Jul 6. Plant Physiol. 2015. PMID: 26149571 Free PMC article. Review.
-
Vernalization: a model for investigating epigenetics and eukaryotic gene regulation in plants.Biochim Biophys Acta. 2007 May-Jun;1769(5-6):269-75. doi: 10.1016/j.bbaexp.2007.02.003. Epub 2007 Feb 27. Biochim Biophys Acta. 2007. PMID: 17383745 Review.
Cited by
-
The Effect of Ambient Temperature on Brachypodium distachyon Development.Front Plant Sci. 2019 Aug 21;10:1011. doi: 10.3389/fpls.2019.01011. eCollection 2019. Front Plant Sci. 2019. PMID: 31497030 Free PMC article.
-
Ambient Temperature-Responsive Mechanisms Coordinate Regulation of Flowering Time.Int J Mol Sci. 2018 Oct 16;19(10):3196. doi: 10.3390/ijms19103196. Int J Mol Sci. 2018. PMID: 30332820 Free PMC article. Review.
-
Changing Responses to Changing Seasons: Natural Variation in the Plasticity of Flowering Time.Plant Physiol. 2017 Jan;173(1):16-26. doi: 10.1104/pp.16.01683. Epub 2016 Nov 21. Plant Physiol. 2017. PMID: 27872243 Free PMC article. Review.
-
Global spatial analysis of Arabidopsis natural variants implicates 5'UTR splicing of LATE ELONGATED HYPOCOTYL in responses to temperature.Plant Cell Environ. 2018 Jul;41(7):1524-1538. doi: 10.1111/pce.13188. Epub 2018 Apr 15. Plant Cell Environ. 2018. PMID: 29520807 Free PMC article.
-
Thermo-Sensitive Alternative Splicing of FLOWERING LOCUS M Is Modulated by Cyclin-Dependent Kinase G2.Front Plant Sci. 2020 Jan 22;10:1680. doi: 10.3389/fpls.2019.01680. eCollection 2019. Front Plant Sci. 2020. PMID: 32038671 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
