

The RNA helicase, eIF4A-1, is required for ovule development and cell size homeostasis in Arabidopsis
- PMID: 26493293
- PMCID: PMC4737287
- DOI: 10.1111/tpj.13062
The RNA helicase, eIF4A-1, is required for ovule development and cell size homeostasis in Arabidopsis
Abstract
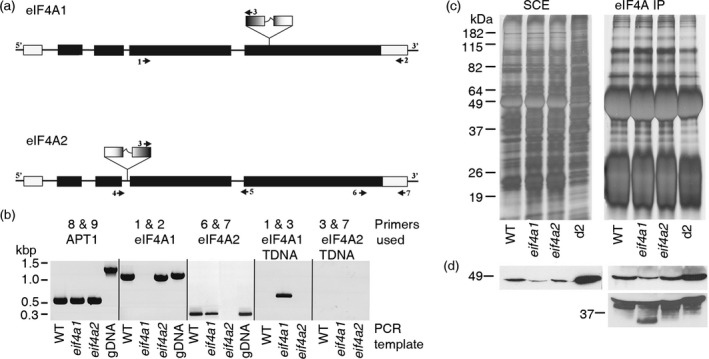
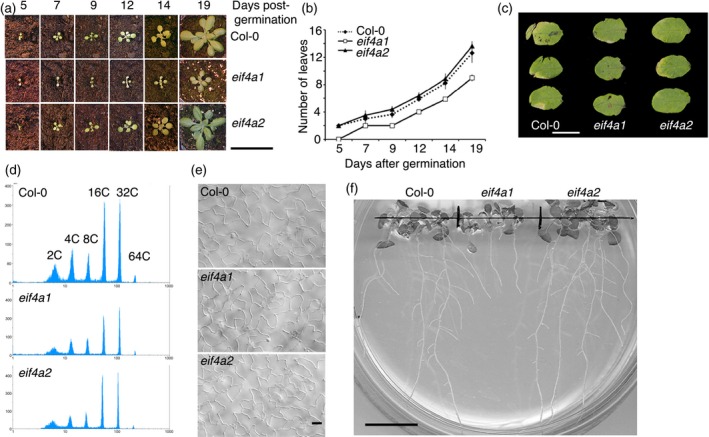
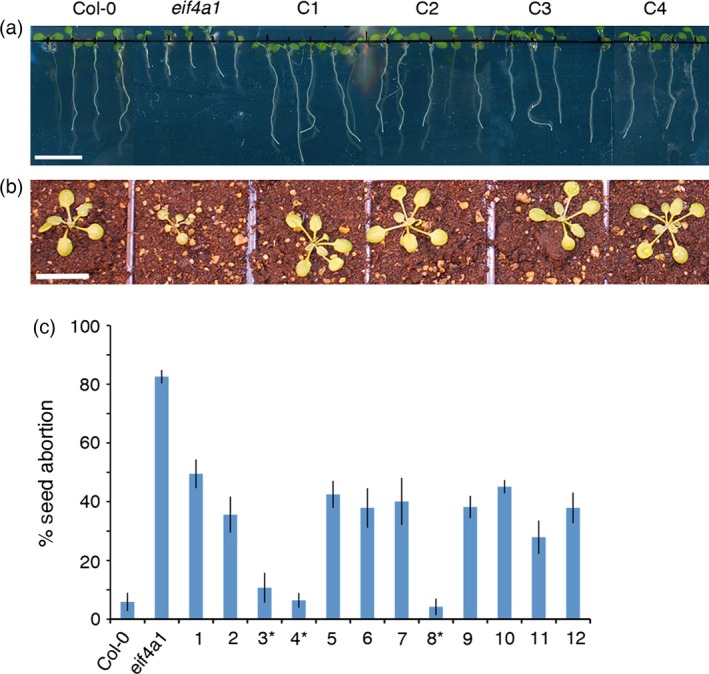
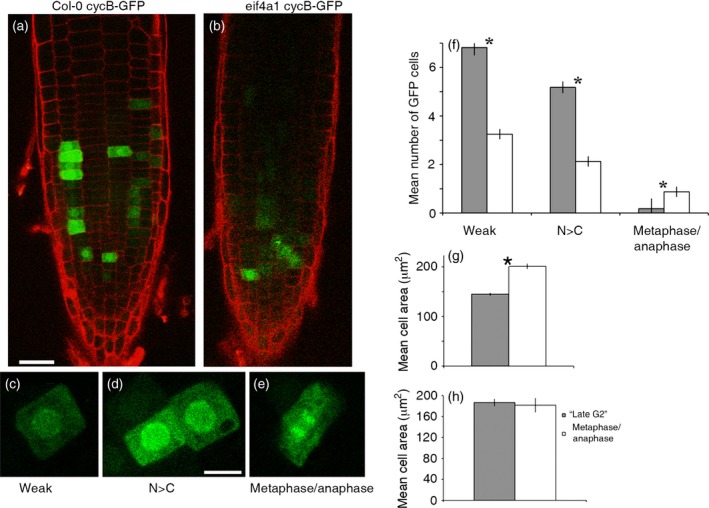
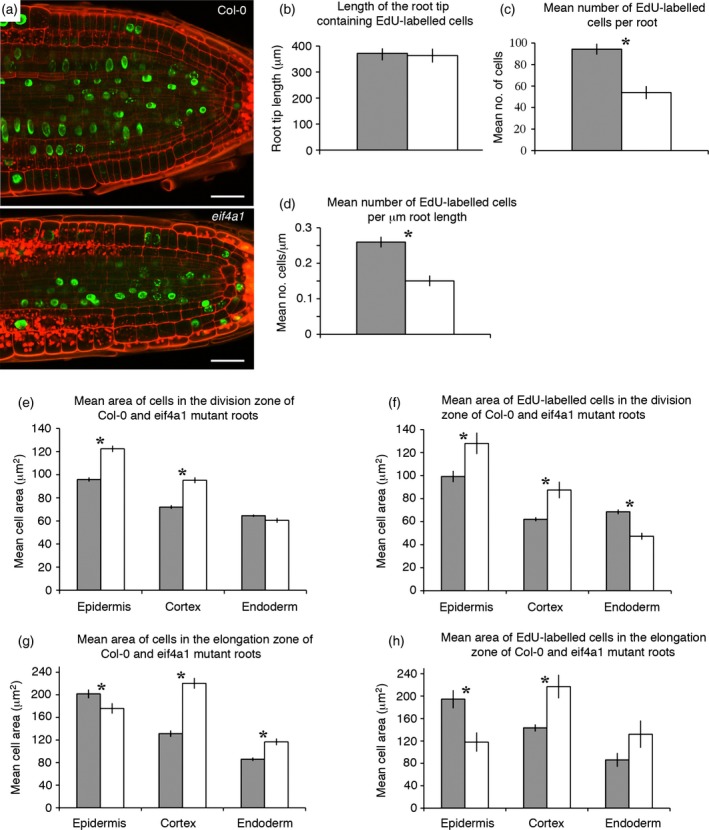
eIF4A is a highly conserved RNA-stimulated ATPase and helicase involved in the initiation of mRNA translation. The Arabidopsis genome encodes two isoforms, one of which (eIF4A-1) is required for the coordination between cell cycle progression and cell size. A T-DNA mutant eif4a1 line, with reduced eIF4A protein levels, displays slow growth, reduced lateral root formation, delayed flowering and abnormal ovule development. Loss of eIF4A-1 reduces the proportion of mitotic cells in the root meristem and perturbs the relationship between cell size and cell cycle progression. Several cell cycle reporter proteins, particularly those expressed at G2/M, have reduced expression in eif4a1 mutant meristems. Single eif4a1 mutants are semisterile and show aberrant ovule growth, whereas double eif4a1 eif4a2 homozygous mutants could not be recovered, indicating that eIF4A function is essential for plant growth and development.
Keywords: Arabidopsis thaliana; DEAD-box helicase; RNA helicase; cell cycle; cell size homeostasis; plant growth; translation factor.
© 2015 The Authors The Plant Journal published by Society for Experimental Biology and John Wiley & Sons Ltd.
Figures

References
-
- Bhat, M. , Robichaud, N. , Hulea, L. , Sonenberg, N. , Pelletier, J. and Topisirovic, I. (2015) Targeting the translation machinery in cancer. Nat. Rev. Drug Discov. 14, 261–278. - PubMed
-
- Bordeleau, M.E. , Cencic, R. , Lindqvist, L. , Oberer, M. , Northcote, P. , Wagner, G. and Pelletier, J. (2006a) RNA‐mediated sequestration of the RNA helicase eIF4A by Pateamine A inhibits translation initiation. Chem. Biol. 13, 1287–1295. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
