

Tubulin cytoskeleton during microsporogenesis in the male-sterile genotype of Allium sativum and fertile Allium ampeloprasum L
- PMID: 26493316
- PMCID: PMC4623095
- DOI: 10.1007/s00497-015-0268-0
Tubulin cytoskeleton during microsporogenesis in the male-sterile genotype of Allium sativum and fertile Allium ampeloprasum L
Abstract
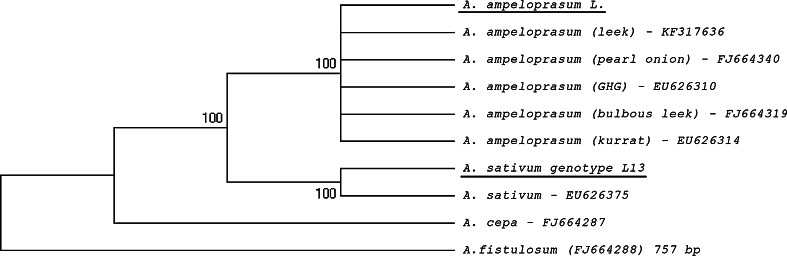
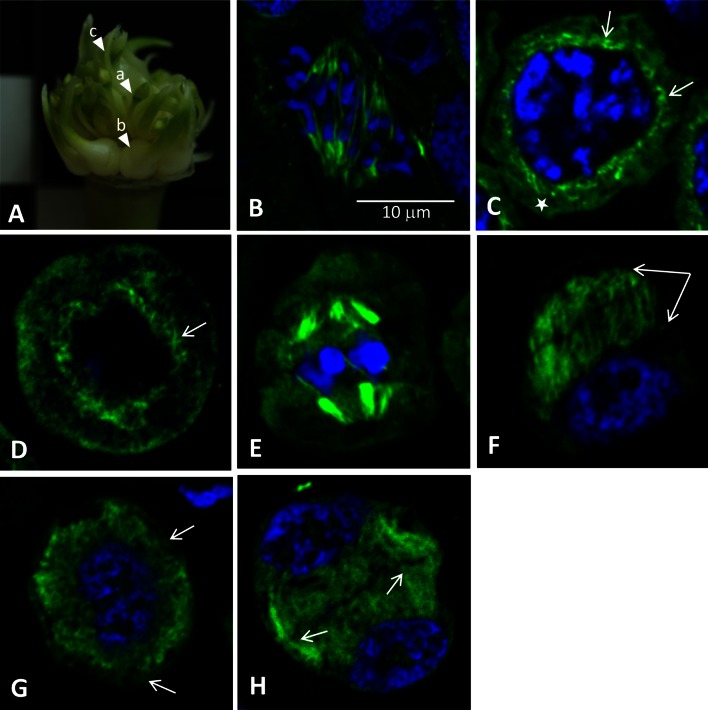
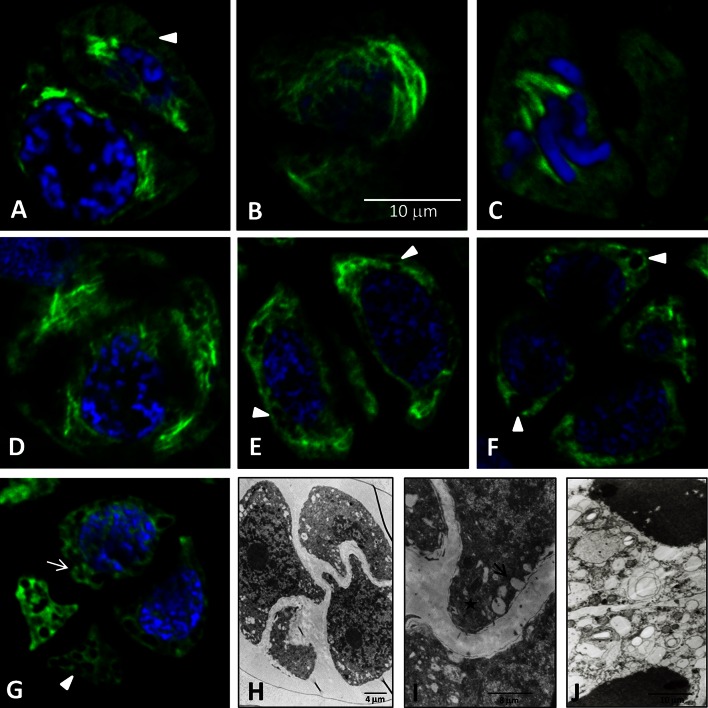
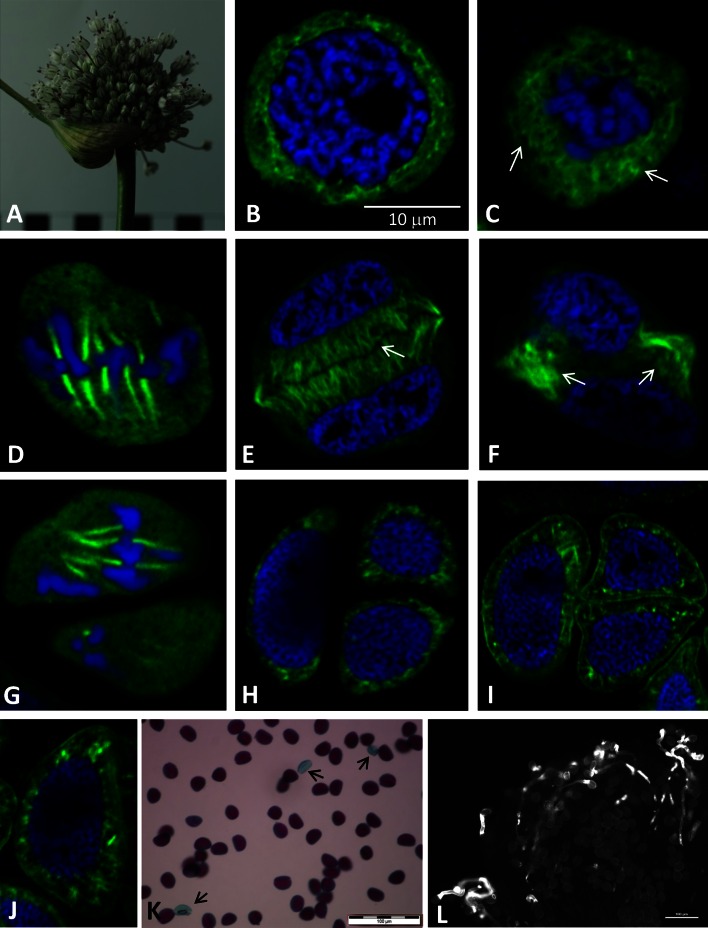
Microsporogenesis in garlic. The male-sterile Allium sativum (garlic) reproduces exclusively in the vegetative mode, and anthropogenic factors seem to be the cause of the loss of sexual reproduction capability. There are many different hypotheses concerning the causes of male sterility in A.sativum; however, the mechanisms underlying this phenomenon have not been comprehensively elucidated.Numerous attempts have been undertaken to understand the causes of male sterility, but the tubulin cytoskeleton in meiotically dividing cells during microsporogenesis has never been investigated in this species. Using sterile A.sativum genotype L13 and its fertile close relative A. ampeloprasum (leek), we have analysed the distribution of the tubulin cytoskeleton during microsporogenesis. We observed that during karyokinesis and cytokinesis, in both meiotic divisions I and II, the microtubular cytoskeleton in garlic L13 formed configurations that resembled tubulin arrangement typical of monocots. However, the tubulin cytoskeleton in garlic was distinctly poorer (composed of a few MT filaments) compared with that found in meiotically dividing cells in A. ampeloprasum. These differences did not affect the course of karyogenesis, chondriokinesis, and cytokinesis, which contributed to completion of microsporogenesis, but there was no further development of the male gametophyte. At the very beginning of the successive stage of development of fertile pollen grains, i.e. gametogenesis, there were disorders involving the absence of a normal cortical cytoskeleton and dramatically progressive degeneration of the cytoplasm in garlic. Therefore,we suggest that, due to disturbances in cortical cytoskeleton formation at the very beginning of gametogenesis, the intracellular transport governed by the cytoskeleton might be perturbed, leading to microspore decay in the male-sterile garlic genotype.
Figures
Similar articles
-
Cytological and biophysical comparative analysis of cell structures at the microsporogenesis stage in sterile and fertile Allium species.Planta. 2017 Jan;245(1):137-150. doi: 10.1007/s00425-016-2597-0. Epub 2016 Sep 29. Planta. 2017. PMID: 27686466 Free PMC article.
-
Characterization of callase (β-1,3-D-glucanase) activity during microsporogenesis in the sterile anthers of Allium sativum L. and the fertile anthers of A. atropurpureum.Sex Plant Reprod. 2012 Jun;25(2):123-31. doi: 10.1007/s00497-012-0184-5. Epub 2012 Mar 23. Sex Plant Reprod. 2012. PMID: 22438078 Free PMC article.
-
Effects of different temperature regimes on flower development, microsporogenesis and fertility in bolting garlic (Allium sativum).Funct Plant Biol. 2015 Jun;42(6):514-526. doi: 10.1071/FP14262. Funct Plant Biol. 2015. PMID: 32480697
-
Chondriokinesis during microsporogenesis in plants.Planta. 2017 Jul;246(1):1-18. doi: 10.1007/s00425-017-2706-8. Epub 2017 May 8. Planta. 2017. PMID: 28484865 Free PMC article. Review.
-
Plants of the genus Allium as antibacterial agents: From tradition to pharmacy.Cell Mol Biol (Noisy-le-grand). 2016 Aug 29;62(9):57-68. Cell Mol Biol (Noisy-le-grand). 2016. PMID: 27585263 Review.
Cited by
-
Cytological and biophysical comparative analysis of cell structures at the microsporogenesis stage in sterile and fertile Allium species.Planta. 2017 Jan;245(1):137-150. doi: 10.1007/s00425-016-2597-0. Epub 2016 Sep 29. Planta. 2017. PMID: 27686466 Free PMC article.
-
MiRNAs differentially expressed in vegetative and reproductive organs of Marchantia polymorpha - insights into their expression pattern, gene structures and function.RNA Biol. 2024 Jan;21(1):1-12. doi: 10.1080/15476286.2024.2303555. Epub 2024 Feb 1. RNA Biol. 2024. PMID: 38303117 Free PMC article.
-
Genome-Wide Identification and Expression Analysis of TUA and TUB Genes in Wheat (Triticum aestivum L.) during Its Development.Plants (Basel). 2022 Dec 13;11(24):3495. doi: 10.3390/plants11243495. Plants (Basel). 2022. PMID: 36559605 Free PMC article.
-
Heteromorphism of stamens in monoclinous flowers of Tinantia erecta (Jacq.) Fenzl as an example of high variability of the androecium in the Commelinaceae family.Protoplasma. 2020 Sep;257(5):1473-1485. doi: 10.1007/s00709-020-01522-z. Epub 2020 Jun 26. Protoplasma. 2020. PMID: 32588232 Free PMC article.
-
Morphological Features of the Anther Development in Tomato Plants with Non-Specific Male Sterility.Biology (Basel). 2020 Feb 17;9(2):32. doi: 10.3390/biology9020032. Biology (Basel). 2020. PMID: 32079211 Free PMC article.
References
-
- Baluska F, Barlow PW. The role of the microtubular cytoskeleton in determining nuclear chromatin structure and passage of maize root cells through the cell cycle. Eur J Cell Biol. 1993;61:160–167. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
