

Molecular architecture of the DED chains at the DISC: regulation of procaspase-8 activation by short DED proteins c-FLIP and procaspase-8 prodomain
- PMID: 26494467
- PMCID: PMC4986640
- DOI: 10.1038/cdd.2015.137
Molecular architecture of the DED chains at the DISC: regulation of procaspase-8 activation by short DED proteins c-FLIP and procaspase-8 prodomain
Abstract
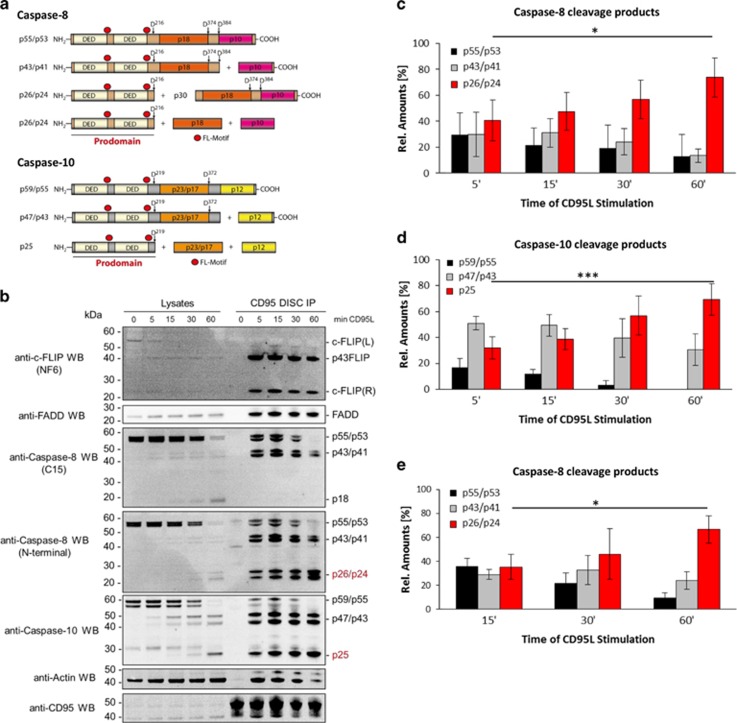
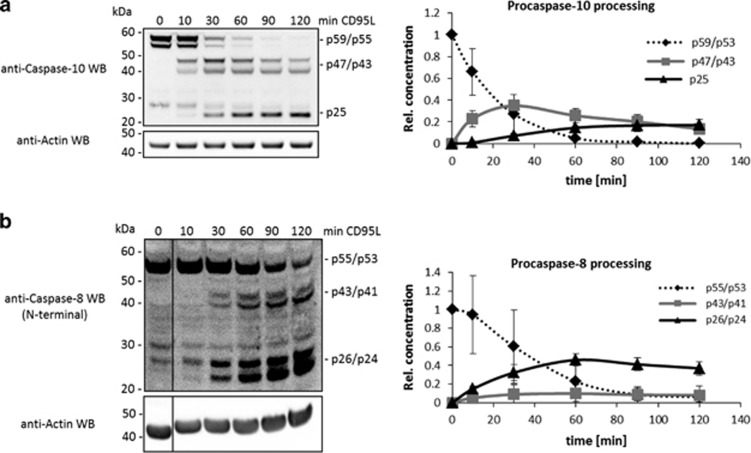
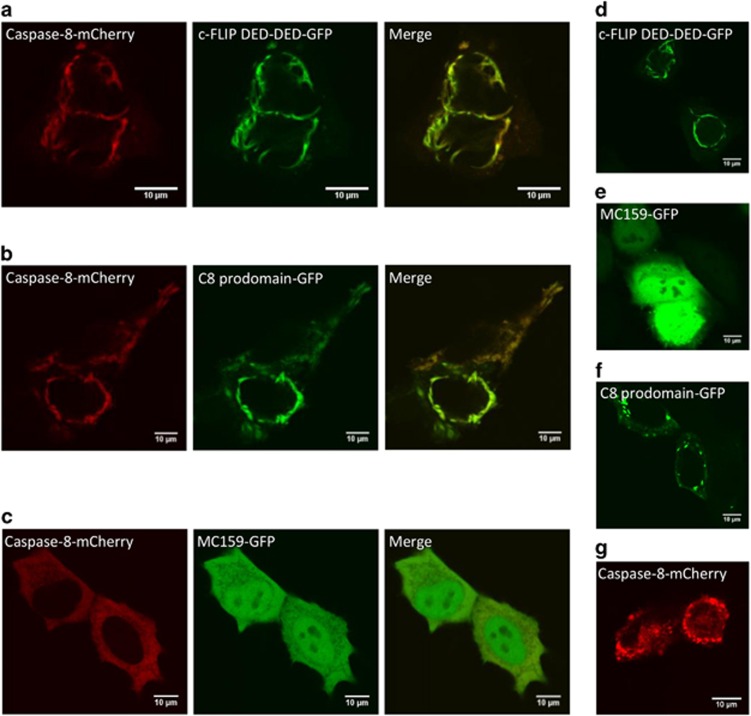
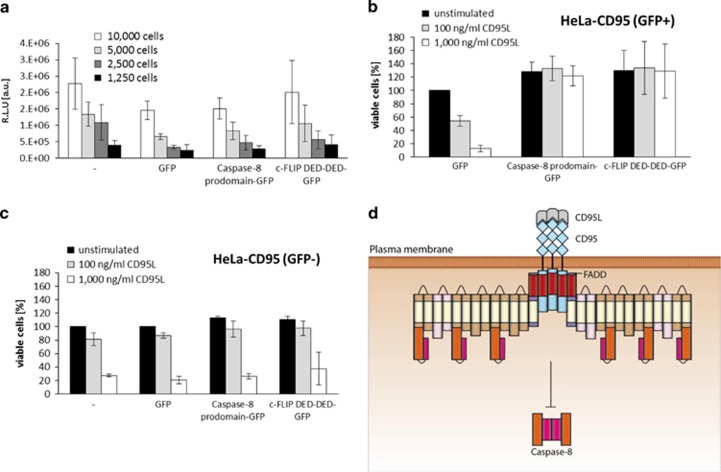
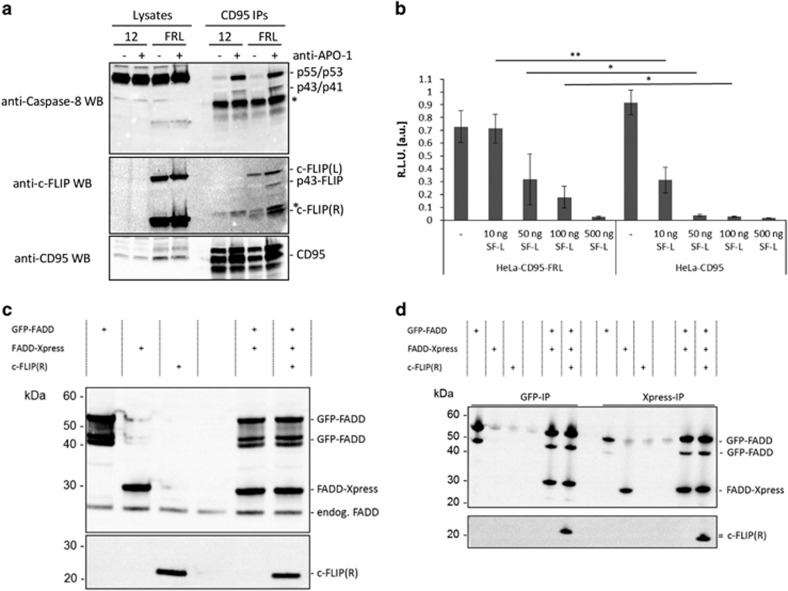
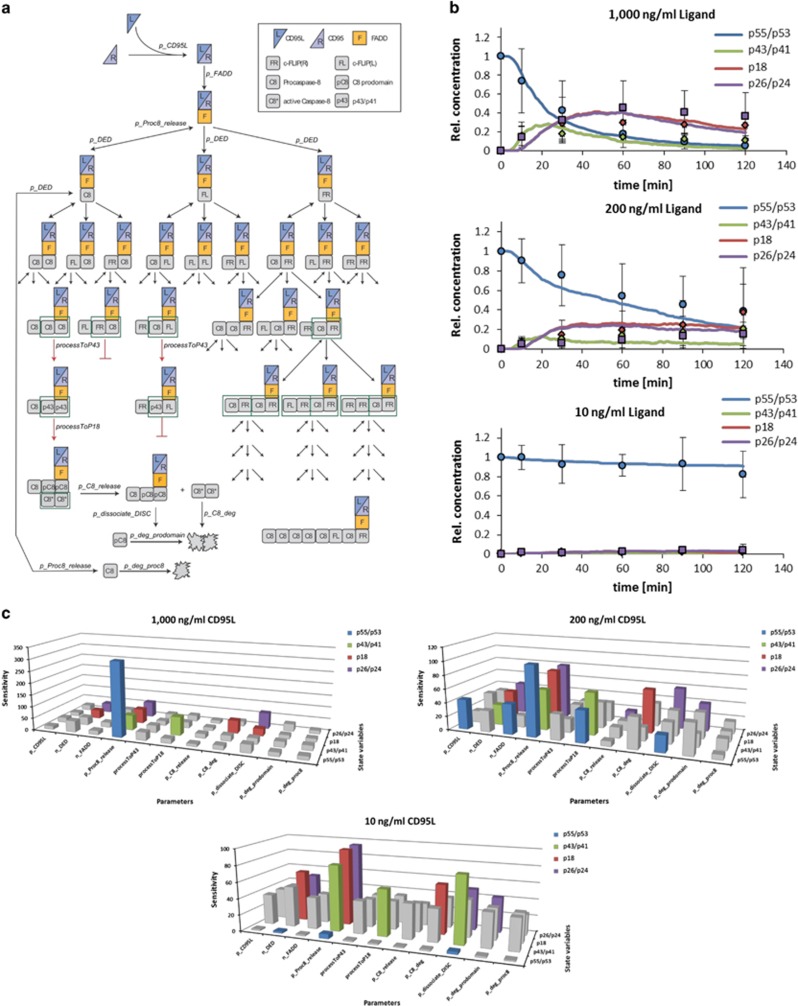
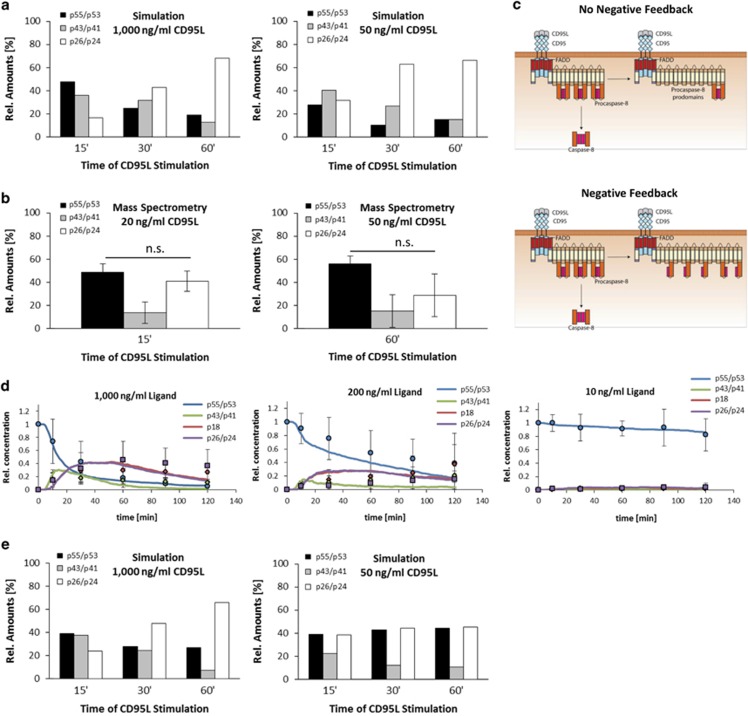
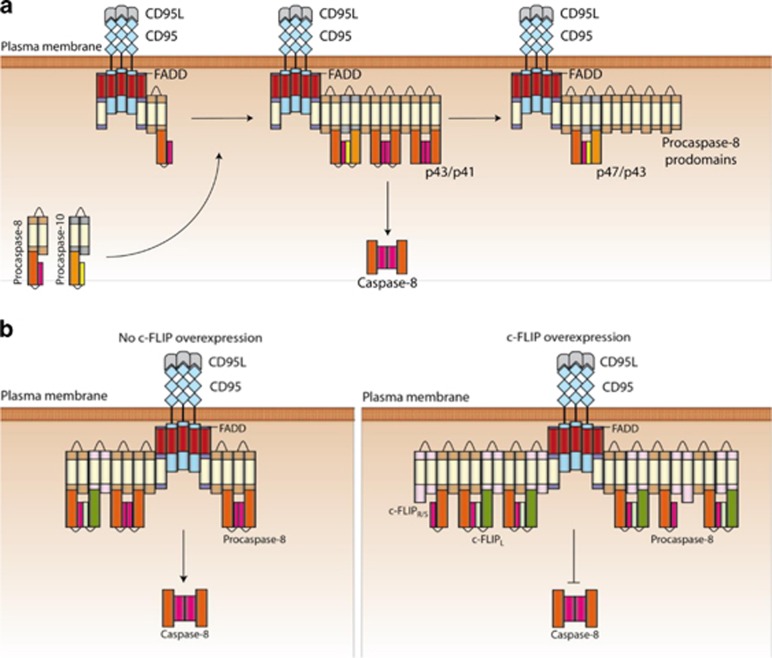
The CD95/Fas/APO-1 death-inducing signaling complex (DISC), comprising CD95, FADD, procaspase-8, procaspase-10, and c-FLIP, has a key role in apoptosis induction. Recently, it was demonstrated that procaspase-8 activation is driven by death effector domain (DED) chains at the DISC. Here, we analyzed the molecular architecture of the chains and the role of the short DED proteins in regulating procaspase-8 activation in the chain model. We demonstrate that the DED chains are largely composed of procaspase-8 cleavage products and, in particular, of its prodomain. The DED chain also comprises c-FLIP and procaspase-10 that are present in 10 times lower amounts compared with procaspase-8. We show that short c-FLIP isoforms can inhibit CD95-induced cell death upon overexpression, likely by forming inactive heterodimers with procaspase-8. Furthermore, we have addressed mechanisms of the termination of chain elongation using experimental and mathematical modeling approaches. We show that neither c-FLIP nor procaspase-8 prodomain terminates the DED chain, but rather the dissociation/association rates of procaspase-8 define the stability of the chain and thereby its length. In addition, we provide evidence that procaspase-8 prodomain generated at the DISC constitutes a negative feedback loop in procaspase-8 activation. Overall, these findings provide new insights into caspase-8 activation in DED chains and apoptosis initiation.
Figures
Similar articles
-
Stoichiometry of the CD95 death-inducing signaling complex: experimental and modeling evidence for a death effector domain chain model.Mol Cell. 2012 Jul 27;47(2):306-19. doi: 10.1016/j.molcel.2012.05.006. Epub 2012 Jun 7. Mol Cell. 2012. PMID: 22683265
-
Long and short isoforms of c-FLIP act as control checkpoints of DED filament assembly.Oncogene. 2020 Feb;39(8):1756-1772. doi: 10.1038/s41388-019-1100-3. Epub 2019 Nov 18. Oncogene. 2020. PMID: 31740779
-
Caspase-8 activity has an essential role in CD95/Fas-mediated MAPK activation.Cell Death Dis. 2011 Oct 6;2(10):e212. doi: 10.1038/cddis.2011.93. Cell Death Dis. 2011. PMID: 21975294 Free PMC article.
-
Regulation of CD95/Fas signaling at the DISC.Cell Death Differ. 2012 Jan;19(1):36-41. doi: 10.1038/cdd.2011.155. Epub 2011 Nov 11. Cell Death Differ. 2012. PMID: 22075988 Free PMC article. Review.
-
Death effecter domain for the assembly of death-inducing signaling complex.Apoptosis. 2015 Feb;20(2):235-9. doi: 10.1007/s10495-014-1060-6. Apoptosis. 2015. PMID: 25451007 Review.
Cited by
-
ITCH-dependent proteasomal degradation of c-FLIP induced by the anti-HER3 antibody 9F7-F11 promotes DR5/caspase 8-mediated apoptosis of tumor cells.Cell Commun Signal. 2019 Aug 23;17(1):106. doi: 10.1186/s12964-019-0413-8. Cell Commun Signal. 2019. PMID: 31443721 Free PMC article.
-
A Comprehensive Exploration of Caspase Detection Methods: From Classical Approaches to Cutting-Edge Innovations.Int J Mol Sci. 2024 May 17;25(10):5460. doi: 10.3390/ijms25105460. Int J Mol Sci. 2024. PMID: 38791499 Free PMC article. Review.
-
Primary and Secondary micro-RNA Modulation the Extrinsic Pathway of Apoptosis in Hepatocellular Carcinoma.Mol Biol. 2023;57(2):165-175. doi: 10.1134/S0026893323020103. Epub 2023 Apr 26. Mol Biol. 2023. PMID: 37128213 Free PMC article.
-
Targeting apoptotic caspases in cancer.Biochim Biophys Acta Mol Cell Res. 2020 Jun;1867(6):118688. doi: 10.1016/j.bbamcr.2020.118688. Epub 2020 Feb 19. Biochim Biophys Acta Mol Cell Res. 2020. PMID: 32087180 Free PMC article. Review.
-
A rare immunological disease, caspase 8 deficiency: case report and literature review.Allergy Asthma Clin Immunol. 2023 Apr 10;19(1):29. doi: 10.1186/s13223-023-00778-3. Allergy Asthma Clin Immunol. 2023. PMID: 37038193 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
