

Silencio/CG9754 connects the Piwi-piRNA complex to the cellular heterochromatin machinery
- PMID: 26494711
- PMCID: PMC4647559
- DOI: 10.1101/gad.271908.115
Silencio/CG9754 connects the Piwi-piRNA complex to the cellular heterochromatin machinery
Abstract
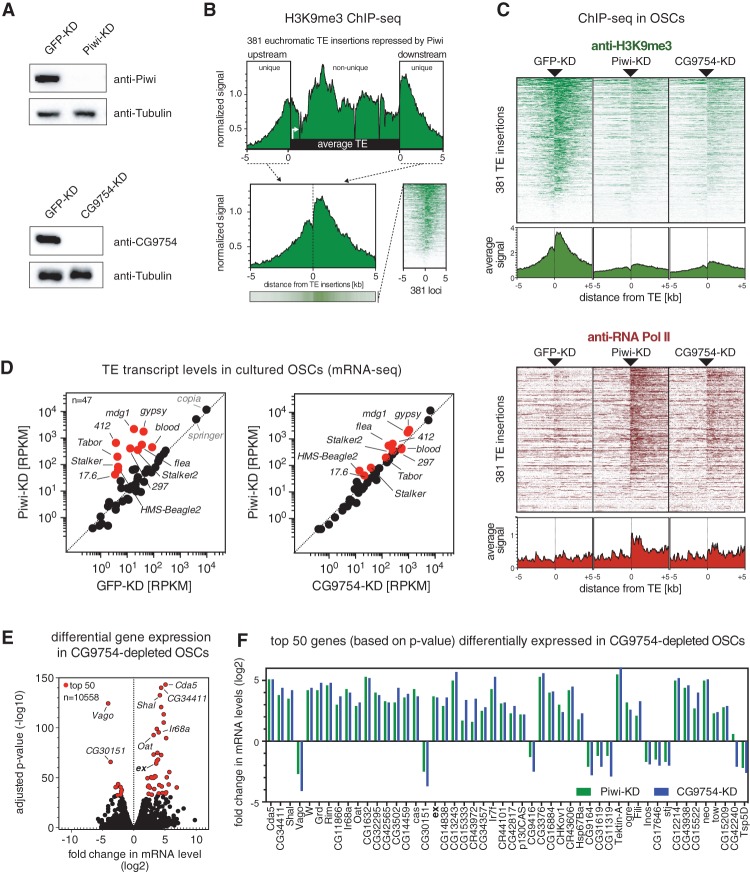
The repression of transposable elements in eukaryotes often involves their transcriptional silencing via targeted chromatin modifications. In animal gonads, nuclear Argonaute proteins of the PIWI clade complexed with small guide RNAs (piRNAs) serve as sequence specificity determinants in this process. How binding of nuclear PIWI-piRNA complexes to nascent transcripts orchestrates heterochromatin formation and transcriptional silencing is unknown. Here, we characterize CG9754/Silencio as an essential piRNA pathway factor that is required for Piwi-mediated transcriptional silencing in Drosophila. Ectopic targeting of Silencio to RNA or DNA is sufficient to elicit silencing independently of Piwi and known piRNA pathway factors. Instead, Silencio requires the H3K9 methyltransferase Eggless/SetDB1 for its silencing ability. In agreement with this, SetDB1, but not Su(var)3-9, is required for Piwi-mediated transcriptional silencing genome-wide. Due to its interaction with the target-engaged Piwi-piRNA complex, we suggest that Silencio acts as linker between the sequence specificity factor Piwi and the cellular heterochromatin machinery.
Keywords: H3K9 methylation; Piwi; heterochromatin formation; piRNA pathway; transcriptional silencing; transposon silencing.
© 2015 Sienski et al.; Published by Cold Spring Harbor Laboratory Press.
Figures





Similar articles
-
Panoramix SUMOylation on chromatin connects the piRNA pathway to the cellular heterochromatin machinery.Nat Struct Mol Biol. 2022 Feb;29(2):130-142. doi: 10.1038/s41594-022-00721-x. Epub 2022 Feb 16. Nat Struct Mol Biol. 2022. PMID: 35173350 Free PMC article.
-
A Pandas complex adapted for piRNA-guided transcriptional silencing and heterochromatin formation.Nat Cell Biol. 2019 Oct;21(10):1261-1272. doi: 10.1038/s41556-019-0396-0. Epub 2019 Sep 30. Nat Cell Biol. 2019. PMID: 31570835
-
piRNA-mediated regulation of transposon alternative splicing in the soma and germ line.Nature. 2017 Dec 14;552(7684):268-272. doi: 10.1038/nature25018. Epub 2017 Dec 6. Nature. 2017. PMID: 29211718 Free PMC article.
-
To export, or not to export: how nuclear export factor variants resolve Piwi's dilemma.Biochem Soc Trans. 2021 Nov 1;49(5):2073-2079. doi: 10.1042/BST20201171. Biochem Soc Trans. 2021. PMID: 34643228 Review.
-
Two distinct transcriptional controls triggered by nuclear Piwi-piRISCs in the Drosophila piRNA pathway.Curr Opin Struct Biol. 2018 Dec;53:69-76. doi: 10.1016/j.sbi.2018.06.005. Epub 2018 Jul 7. Curr Opin Struct Biol. 2018. PMID: 29990672 Review.
Cited by
-
Restricting retrotransposons: a review.Mob DNA. 2016 Aug 11;7:16. doi: 10.1186/s13100-016-0070-z. eCollection 2016. Mob DNA. 2016. PMID: 27525044 Free PMC article. Review.
-
Su(var)2-10 and the SUMO Pathway Link piRNA-Guided Target Recognition to Chromatin Silencing.Mol Cell. 2020 Feb 6;77(3):556-570.e6. doi: 10.1016/j.molcel.2019.11.012. Epub 2019 Dec 31. Mol Cell. 2020. PMID: 31901446 Free PMC article.
-
The Mi-2 nucleosome remodeler and the Rpd3 histone deacetylase are involved in piRNA-guided heterochromatin formation.Nat Commun. 2020 Jun 4;11(1):2818. doi: 10.1038/s41467-020-16635-5. Nat Commun. 2020. PMID: 32499524 Free PMC article.
-
TEtools facilitates big data expression analysis of transposable elements and reveals an antagonism between their activity and that of piRNA genes.Nucleic Acids Res. 2017 Feb 28;45(4):e17. doi: 10.1093/nar/gkw953. Nucleic Acids Res. 2017. PMID: 28204592 Free PMC article.
-
RNA matchmaking in chromatin regulation.Biochem Soc Trans. 2020 Dec 18;48(6):2467-2481. doi: 10.1042/BST20191225. Biochem Soc Trans. 2020. PMID: 33245317 Free PMC article. Review.
References
-
- Ayyanathan K, Lechner MS, Bell P, Maul GG, Schultz DC, Yamada Y, Tanaka K, Torigoe K, Rauscher FJ III. 2003. Regulated recruitment of HP1 to a euchromatic gene induces mitotically heritable, epigenetic gene silencing: a mammalian cell culture model of gene variegation. Genes Dev 17: 1855–1869. - PMC - PubMed
-
- Baron-Benhamou J, Gehring NH, Kulozik AE, Hentze MW. 2004. Using the λN peptide to tether proteins to RNAs. Methods Mol Biol 257: 135–154. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases