

FoxP1 orchestration of ASD-relevant signaling pathways in the striatum
- PMID: 26494785
- PMCID: PMC4617974
- DOI: 10.1101/gad.267989.115
FoxP1 orchestration of ASD-relevant signaling pathways in the striatum
Abstract
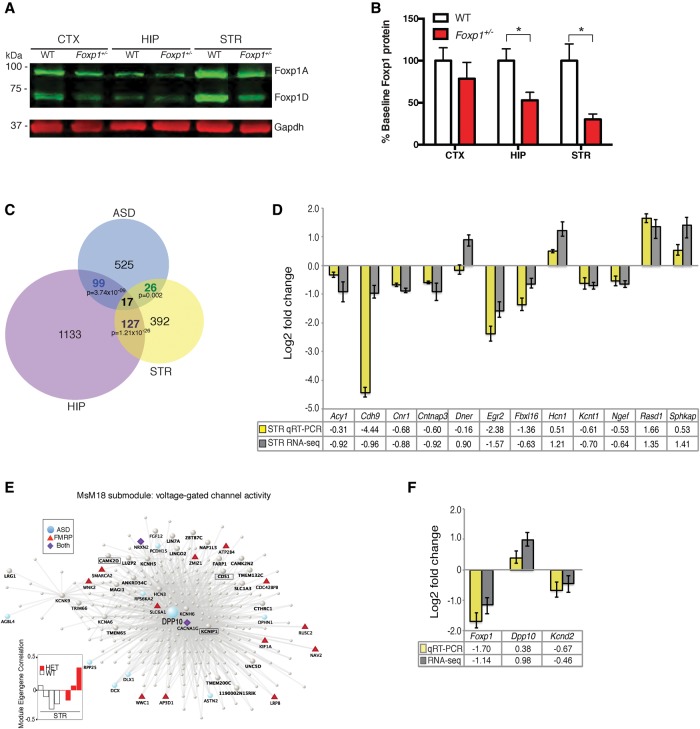
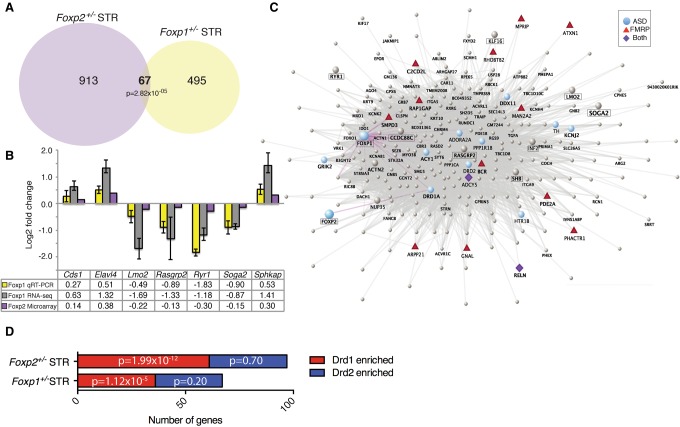
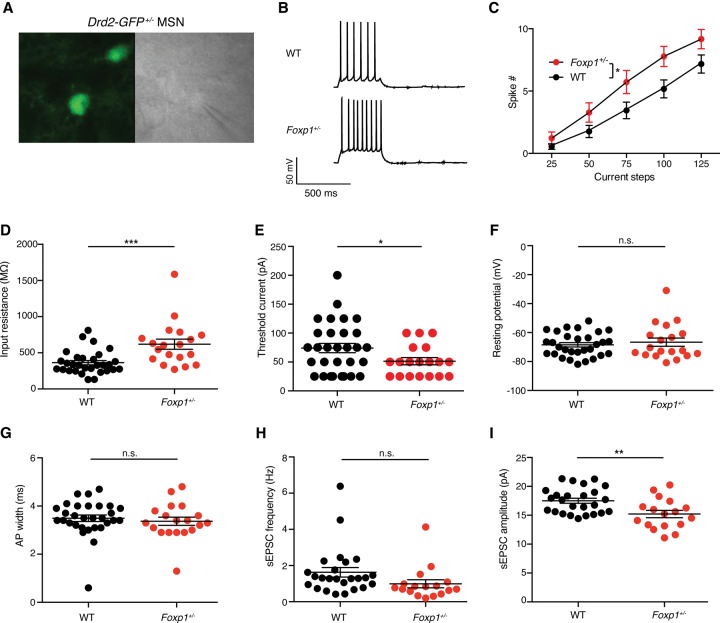
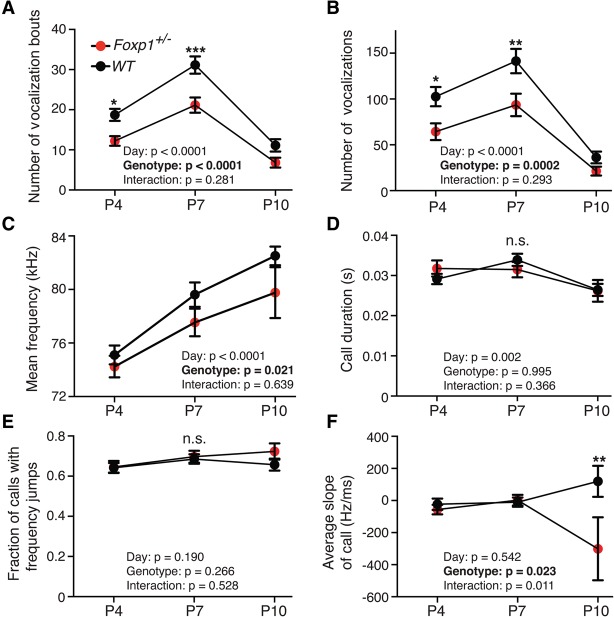
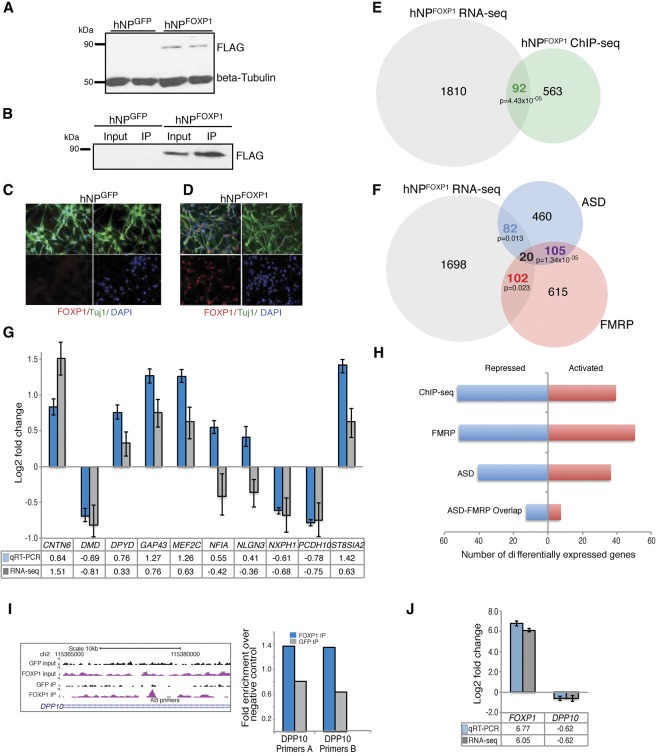
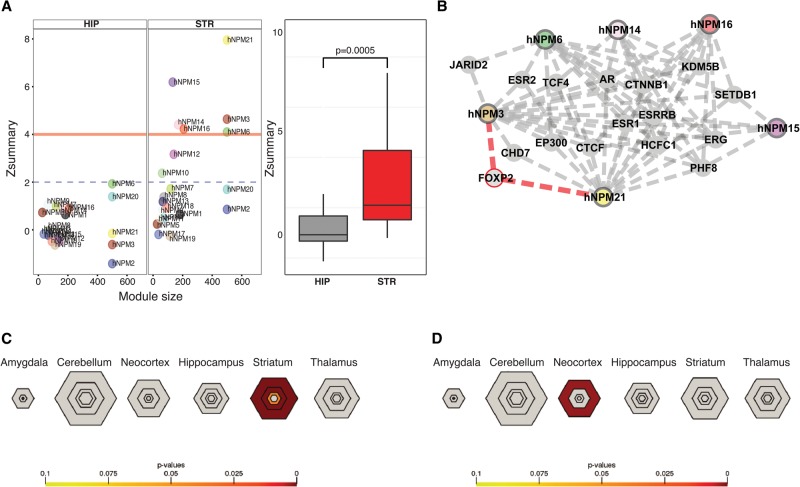
Mutations in the transcription factor Forkhead box p1 (FOXP1) are causative for neurodevelopmental disorders such as autism. However, the function of FOXP1 within the brain remains largely uncharacterized. Here, we identify the gene expression program regulated by FoxP1 in both human neural cells and patient-relevant heterozygous Foxp1 mouse brains. We demonstrate a role for FoxP1 in the transcriptional regulation of autism-related pathways as well as genes involved in neuronal activity. We show that Foxp1 regulates the excitability of striatal medium spiny neurons and that reduction of Foxp1 correlates with defects in ultrasonic vocalizations. Finally, we demonstrate that FoxP1 has an evolutionarily conserved role in regulating pathways involved in striatal neuron identity through gene expression studies in human neural progenitors with altered FOXP1 levels. These data support an integral role for FoxP1 in regulating signaling pathways vulnerable in autism and the specific regulation of striatal pathways important for vocal communication.
Keywords: autism; gene expression; neuronal activity; striatum; ultrasonic vocalizations.
© 2015 Araujo et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
-
- Agmon A, Connors BW. 1991. Thalamocortical responses of mouse somatosensory (barrel) cortex in vitro. Neuroscience 41: 365–379. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases