

Constitutive BAK activation as a determinant of drug sensitivity in malignant lymphohematopoietic cells
- PMID: 26494789
- PMCID: PMC4617978
- DOI: 10.1101/gad.267997.115
Constitutive BAK activation as a determinant of drug sensitivity in malignant lymphohematopoietic cells
Abstract
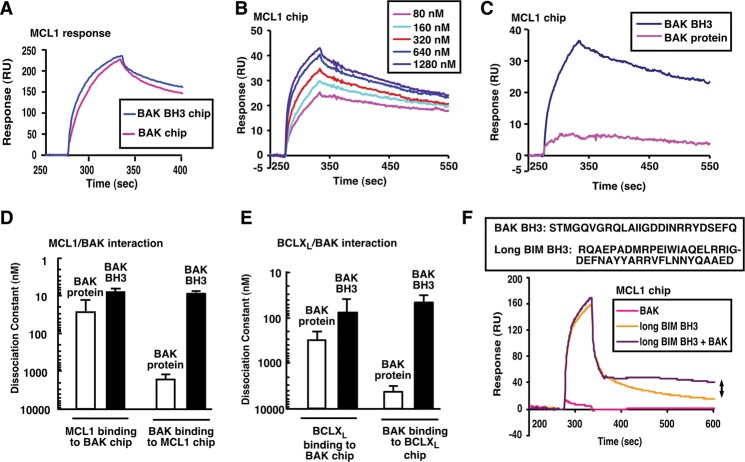
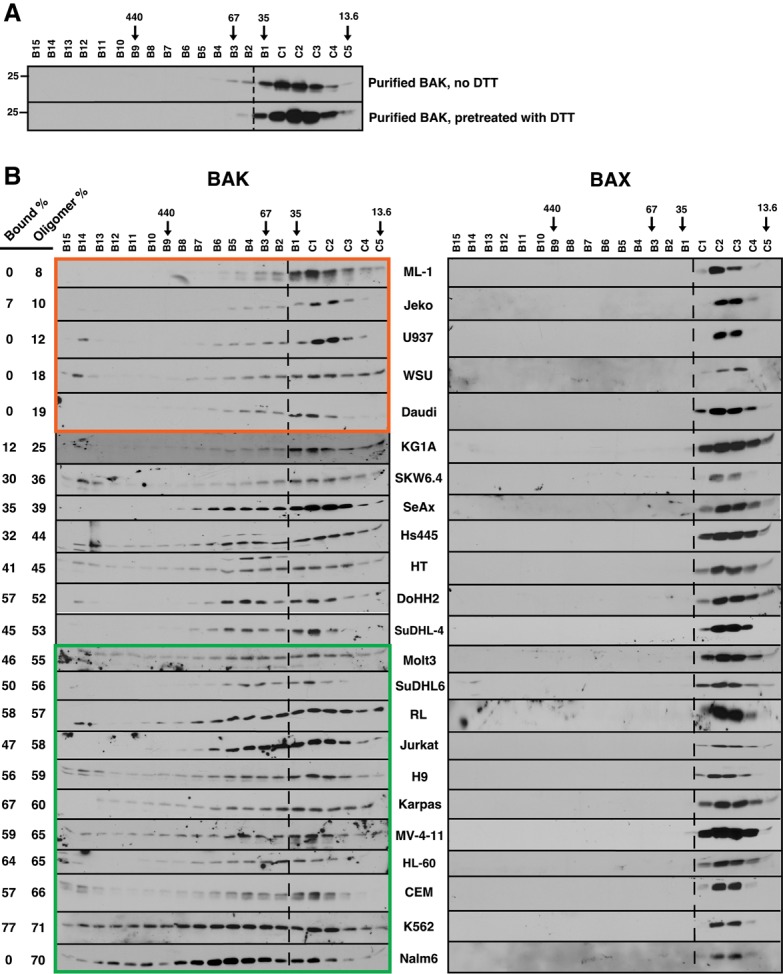
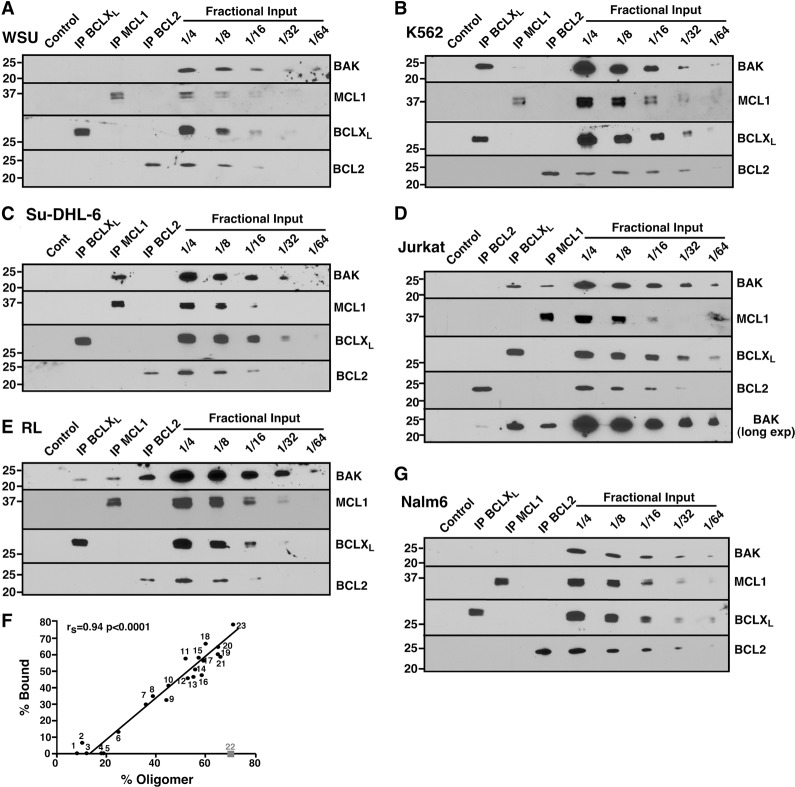
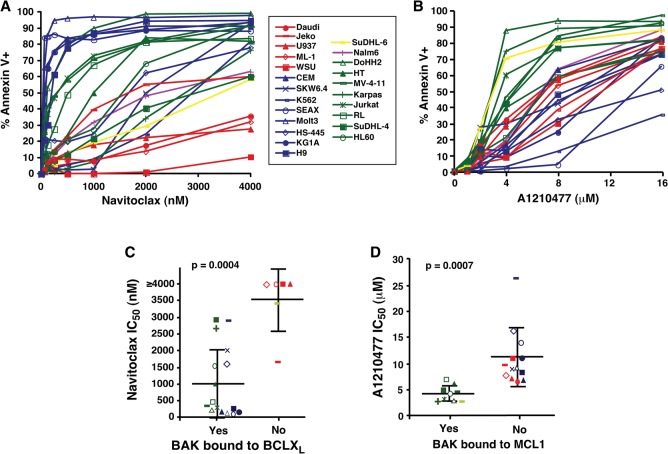
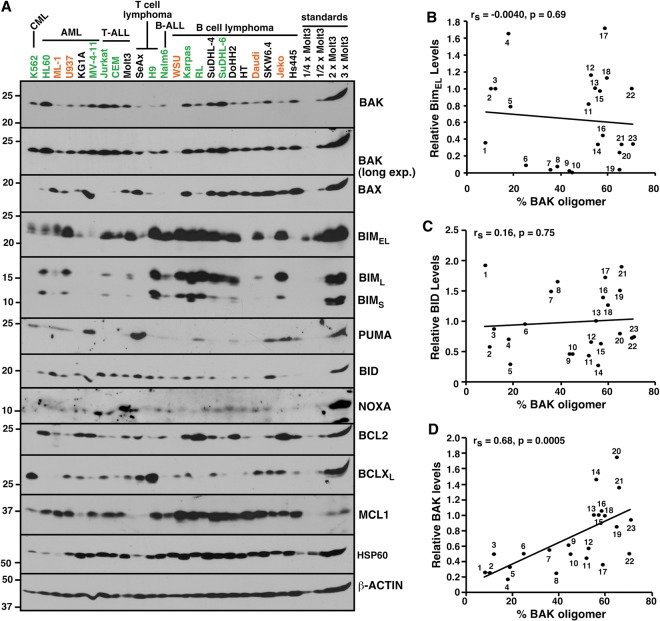
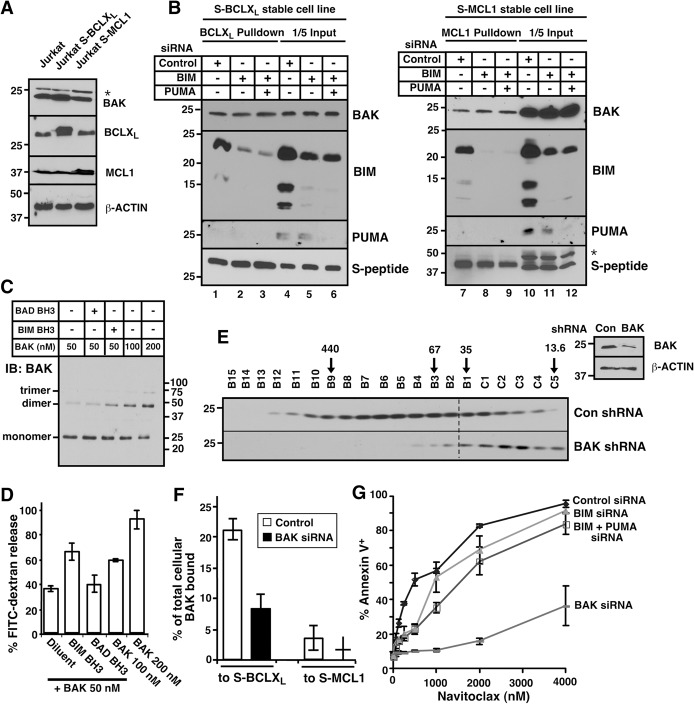
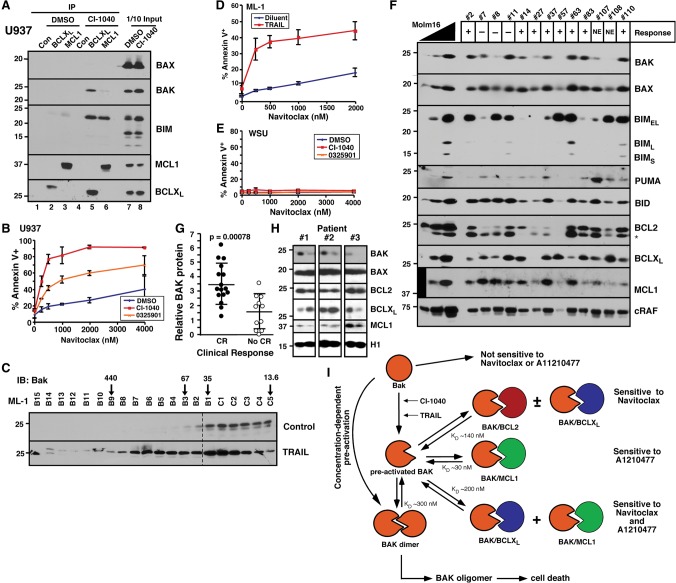
Mitochondrial outer membrane permeabilization (MOMP), a key step in the intrinsic apoptotic pathway, is incompletely understood. Current models emphasize the role of BH3-only BCL2 family members in BAX and BAK activation. Here we demonstrate concentration-dependent BAK autoactivation under cell-free conditions and provide evidence that this autoactivation plays a key role in regulating the intrinsic apoptotic pathway in intact cells. In particular, we show that up to 80% of BAK (but not BAX) in lymphohematopoietic cell lines is oligomerized and bound to anti-apoptotic BCL2 family members in the absence of exogenous death stimuli. The extent of this constitutive BAK oligomerization is diminished by BAK knockdown and unaffected by BIM or PUMA down-regulation. Further analysis indicates that sensitivity of cells to BH3 mimetics reflects the identity of the anti-apoptotic proteins to which BAK is constitutively bound, with extensive BCLXL•BAK complexes predicting navitoclax sensitivity, and extensive MCL1•BAK complexes predicting A1210477 sensitivity. Moreover, high BAK expression correlates with sensitivity of clinical acute myelogenous leukemia to chemotherapy, whereas low BAK levels correlate with resistance and relapse. Collectively, these results inform current understanding of MOMP and provide new insight into the ability of BH3 mimetics to induce apoptosis without directly activating BAX or BAK.
Keywords: BAK; BH3 mimetic; apoptosis.
© 2015 Dai et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
Similar articles
-
Bax/Bak activation in the absence of Bid, Bim, Puma, and p53.Cell Death Dis. 2016 Jun 16;7(6):e2266. doi: 10.1038/cddis.2016.167. Cell Death Dis. 2016. PMID: 27310874 Free PMC article.
-
Cisplatin-induced apoptosis in non-small-cell lung cancer cells is dependent on Bax- and Bak-induction pathway and synergistically activated by BH3-mimetic ABT-263 in p53 wild-type and mutant cells.Biochem Biophys Res Commun. 2016 Apr 29;473(2):490-6. doi: 10.1016/j.bbrc.2016.03.053. Epub 2016 Mar 18. Biochem Biophys Res Commun. 2016. PMID: 26996126
-
BH3-mimetic toolkit guides the respective use of BCL2 and MCL1 BH3-mimetics in myeloma treatment.Blood. 2018 Dec 20;132(25):2656-2669. doi: 10.1182/blood-2018-03-836718. Epub 2018 Oct 11. Blood. 2018. PMID: 30309889
-
BH3 Mimetics for the Treatment of B-Cell Malignancies-Insights and Lessons from the Clinic.Cancers (Basel). 2020 Nov 12;12(11):3353. doi: 10.3390/cancers12113353. Cancers (Basel). 2020. PMID: 33198338 Free PMC article. Review.
-
BCL2 and MCL1 inhibitors for hematologic malignancies.Blood. 2021 Sep 30;138(13):1120-1136. doi: 10.1182/blood.2020006785. Blood. 2021. PMID: 34320168 Review.
Cited by
-
Measurement of BH3-only protein tolerance.Cell Death Differ. 2018 Feb;25(2):282-293. doi: 10.1038/cdd.2017.156. Epub 2017 Oct 20. Cell Death Differ. 2018. PMID: 29053140 Free PMC article.
-
Deciphering molecular specificity in MCL-1/BAK interaction and its implications for designing potent MCL-1 inhibitors.Cell Death Differ. 2025 Jun;32(6):991-999. doi: 10.1038/s41418-025-01454-2. Epub 2025 Feb 3. Cell Death Differ. 2025. PMID: 39901037
-
Characterization of an alternative BAK-binding site for BH3 peptides.Nat Commun. 2020 Jul 3;11(1):3301. doi: 10.1038/s41467-020-17074-y. Nat Commun. 2020. PMID: 32620849 Free PMC article.
-
BAX, BAK, and BOK: A Coming of Age for the BCL-2 Family Effector Proteins.Cold Spring Harb Perspect Biol. 2020 Apr 1;12(4):a036319. doi: 10.1101/cshperspect.a036319. Cold Spring Harb Perspect Biol. 2020. PMID: 31570337 Free PMC article. Review.
-
miR-27a regulates the sensitivity of breast cancer cells to cisplatin treatment via BAK-SMAC/DIABLO-XIAP axis.Tumour Biol. 2016 May;37(5):6837-45. doi: 10.1007/s13277-015-4500-1. Epub 2015 Dec 10. Tumour Biol. 2016. PMID: 26662313
References
-
- Antonsson B, Montessuit S, Sanchez B, Martinou JC. 2001. Bax is present as a high molecular weight oligomer/complex in the mitochondrial membrane of apoptotic cells. J Biol Chem 276: 11615–11623. - PubMed
-
- Brouwer JM, Westphal D, Dewson G, Robin AY, Uren RT, Bartolo R, Thompson GV, Colman PM, Kluck RM, Czabotar PE. 2014. Bak core and latch domains separate during activation, and freed core domains form symmetric homodimers. Mol Cell 55: 938–946. - PubMed
-
- Certo M, Moore Vdel G, Nishino M, Wei G, Korsmeyer S, Armstrong SA, Letai A. 2006. Mitochondria primed by death signals determine cellular addiction to antiapoptotic BCL-2 family members. Cancer Cell 9: 351–365. - PubMed
-
- Cheng EH, Sheiko TV, Fisher JK, Craigen WJ, Korsmeyer SJ. 2003. VDAC2 inhibits BAK activation and mitochondrial apoptosis. Science 301: 513–517. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials