

HENMT1 and piRNA Stability Are Required for Adult Male Germ Cell Transposon Repression and to Define the Spermatogenic Program in the Mouse
- PMID: 26496356
- PMCID: PMC4619860
- DOI: 10.1371/journal.pgen.1005620
HENMT1 and piRNA Stability Are Required for Adult Male Germ Cell Transposon Repression and to Define the Spermatogenic Program in the Mouse
Erratum in
-
Correction: HENMT1 and piRNA Stability Are Required for Adult Male Germ Cell Transposon Repression and to Define the Spermatogenic Program in the Mouse.PLoS Genet. 2015 Dec 29;11(12):e1005782. doi: 10.1371/journal.pgen.1005782. eCollection 2015 Dec. PLoS Genet. 2015. PMID: 26714033 Free PMC article. No abstract available.
Abstract
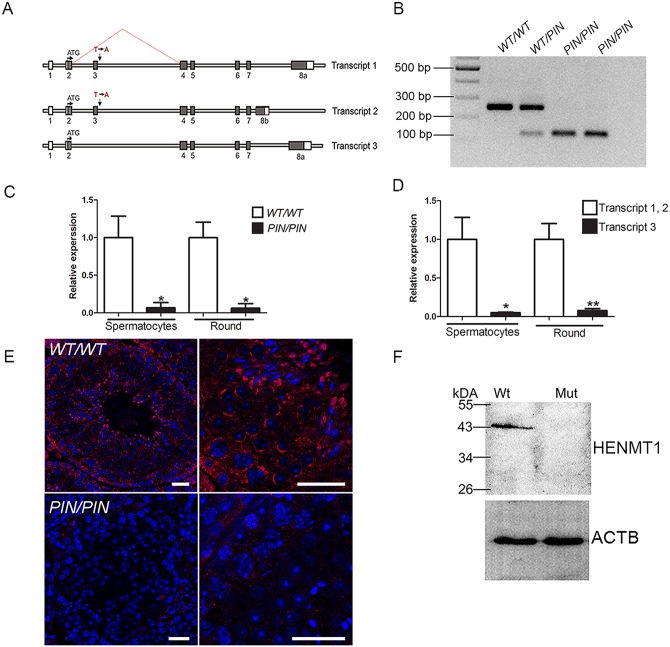
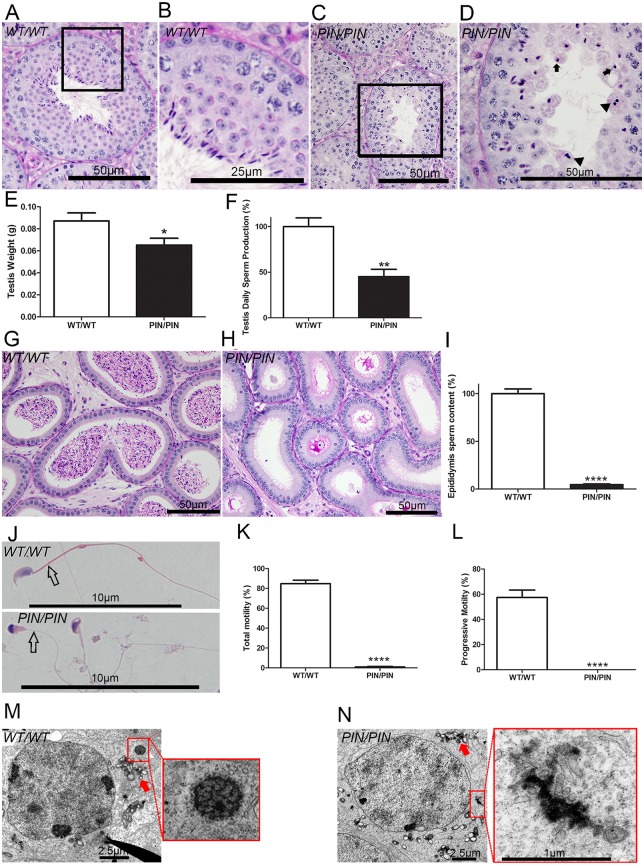
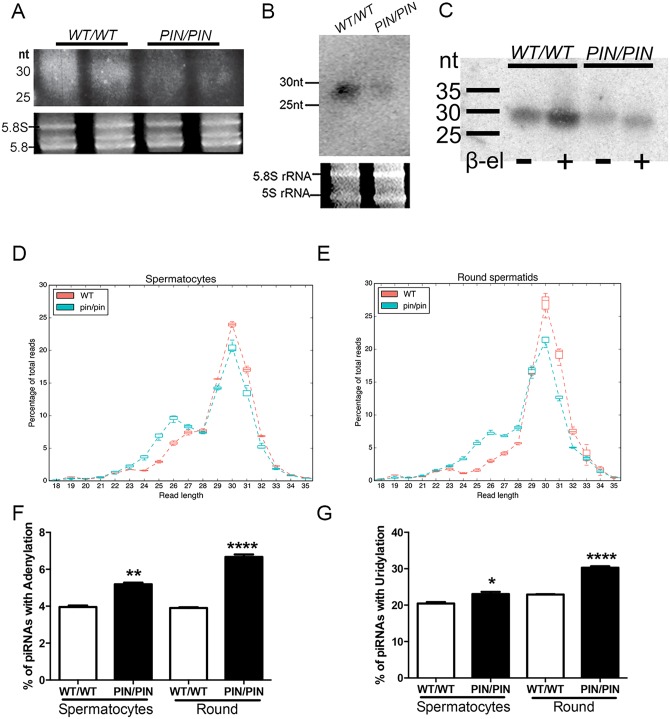
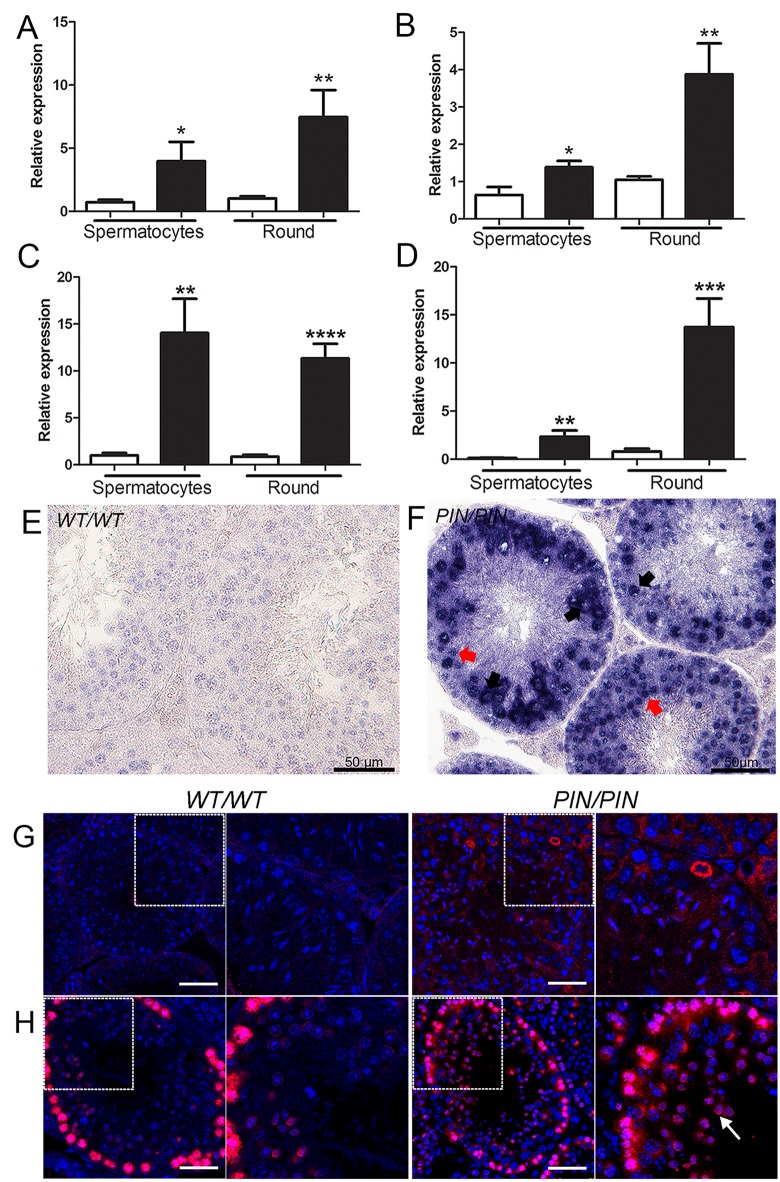
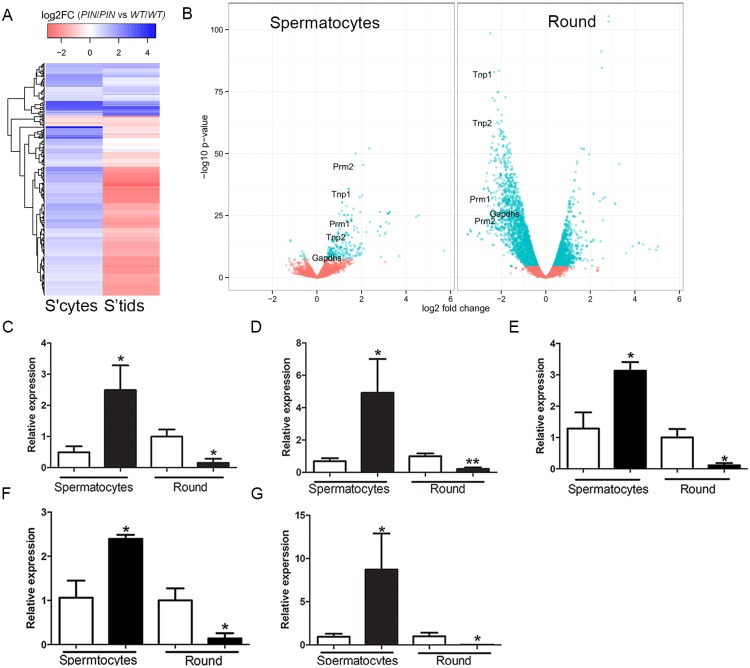
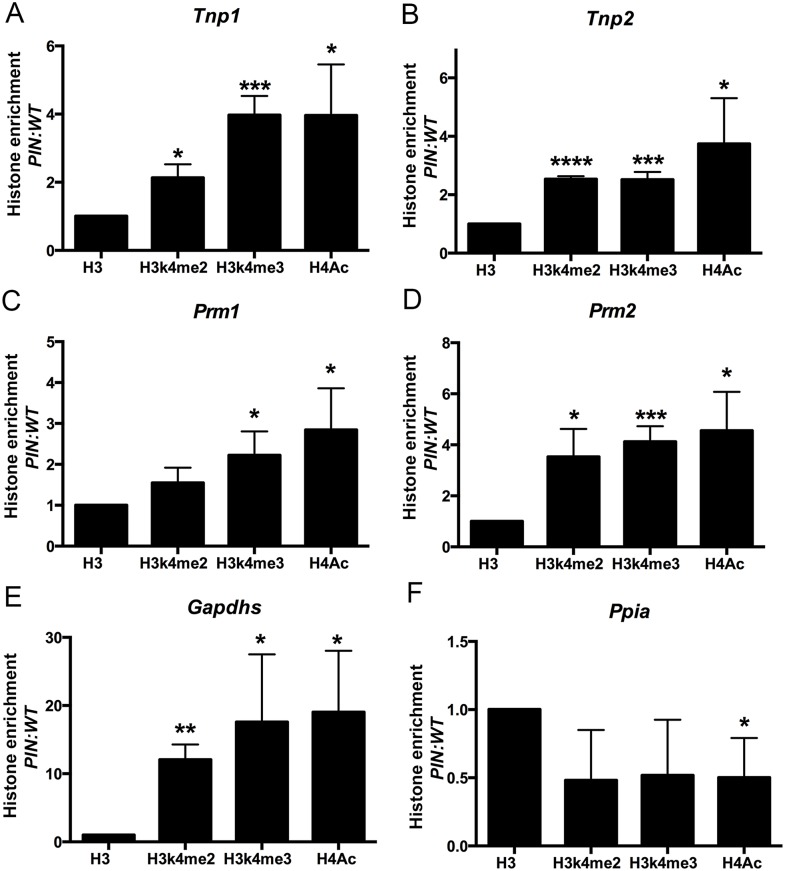
piRNAs are critical for transposable element (TE) repression and germ cell survival during the early phases of spermatogenesis, however, their role in adult germ cells and the relative importance of piRNA methylation is poorly defined in mammals. Using a mouse model of HEN methyltransferase 1 (HENMT1) loss-of-function, RNA-Seq and a range of RNA assays we show that HENMT1 is required for the 2' O-methylation of mammalian piRNAs. HENMT1 loss leads to piRNA instability, reduced piRNA bulk and length, and ultimately male sterility characterized by a germ cell arrest at the elongating germ cell phase of spermatogenesis. HENMT1 loss-of-function, and the concomitant loss of piRNAs, resulted in TE de-repression in adult meiotic and haploid germ cells, and the precocious, and selective, expression of many haploid-transcripts in meiotic cells. Precocious expression was associated with a more active chromatin state in meiotic cells, elevated levels of DNA damage and a catastrophic deregulation of the haploid germ cell gene expression. Collectively these results define a critical role for HENMT1 and piRNAs in the maintenance of TE repression in adult germ cells and setting the spermatogenic program.
Conflict of interest statement
The authors have declared that no competing interests exist
Figures
Similar articles
-
Expression patterns of HENMT1 and PIWIL1 in human testis: implications for transposon expression.Reproduction. 2017 Oct;154(4):363-374. doi: 10.1530/REP-16-0586. Epub 2017 Jul 4. Reproduction. 2017. PMID: 28676534
-
piRNA-directed cleavage of meiotic transcripts regulates spermatogenesis.Genes Dev. 2015 May 15;29(10):1032-44. doi: 10.1101/gad.260455.115. Genes Dev. 2015. PMID: 25995188 Free PMC article.
-
yama, a mutant allele of Mov10l1, disrupts retrotransposon silencing and piRNA biogenesis.PLoS Genet. 2021 Feb 26;17(2):e1009265. doi: 10.1371/journal.pgen.1009265. eCollection 2021 Feb. PLoS Genet. 2021. PMID: 33635934 Free PMC article.
-
[The piRNA pathway and spermatogenesis: Advances in studies].Zhonghua Nan Ke Xue. 2016 Oct;22(10):923-927. Zhonghua Nan Ke Xue. 2016. PMID: 29278475 Review. Chinese.
-
[PIWI/piRNA complex-mediated regulation of spermatogenesis].Zhonghua Nan Ke Xue. 2021 Mar;27(3):262-268. Zhonghua Nan Ke Xue. 2021. PMID: 34914310 Review. Chinese.
Cited by
-
The methyltransferase HEN1 is required in Nematostella vectensis for microRNA and piRNA stability as well as larval metamorphosis.PLoS Genet. 2018 Aug 17;14(8):e1007590. doi: 10.1371/journal.pgen.1007590. eCollection 2018 Aug. PLoS Genet. 2018. PMID: 30118479 Free PMC article.
-
PIWI-interacting RNAs are aberrantly expressed and may serve as novel biomarkers for diagnosis of lung adenocarcinoma.Thorac Cancer. 2021 Sep;12(18):2468-2477. doi: 10.1111/1759-7714.14094. Epub 2021 Aug 3. Thorac Cancer. 2021. PMID: 34346164 Free PMC article.
-
The emergence of piRNAs against transposon invasion to preserve mammalian genome integrity.Nat Commun. 2017 Nov 10;8(1):1411. doi: 10.1038/s41467-017-01049-7. Nat Commun. 2017. PMID: 29127279 Free PMC article. Review.
-
Post-transcriptional regulation in spermatogenesis: all RNA pathways lead to healthy sperm.Cell Mol Life Sci. 2021 Dec;78(24):8049-8071. doi: 10.1007/s00018-021-04012-4. Epub 2021 Nov 8. Cell Mol Life Sci. 2021. PMID: 34748024 Free PMC article. Review.
-
RNA 2'-O-Methylation (Nm) Modification in Human Diseases.Genes (Basel). 2019 Feb 5;10(2):117. doi: 10.3390/genes10020117. Genes (Basel). 2019. PMID: 30764532 Free PMC article. Review.
References
-
- Kazazian HH Jr. (2004) Mobile elements: drivers of genome evolution. Science 303: 1626–1632. - PubMed
-
- Kirino Y, Mourelatos Z (2007) Mouse Piwi-interacting RNAs are 2'-O-methylated at their 3' termini. Nat Struct Mol Biol 14: 347–348. - PubMed
-
- Aravin A, Gaidatzis D, Pfeffer S, Lagos-Quintana M, Landgraf P, et al. (2006) A novel class of small RNAs bind to MILI protein in mouse testes. Nature 442: 203–207. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
